 Catalogue PIGMA
Catalogue PIGMA
2023
Type of resources
Available actions
Topics
Keywords
Contact for the resource
Provided by
Years
Formats
Representation types
Update frequencies
status
Scale
Resolution
-
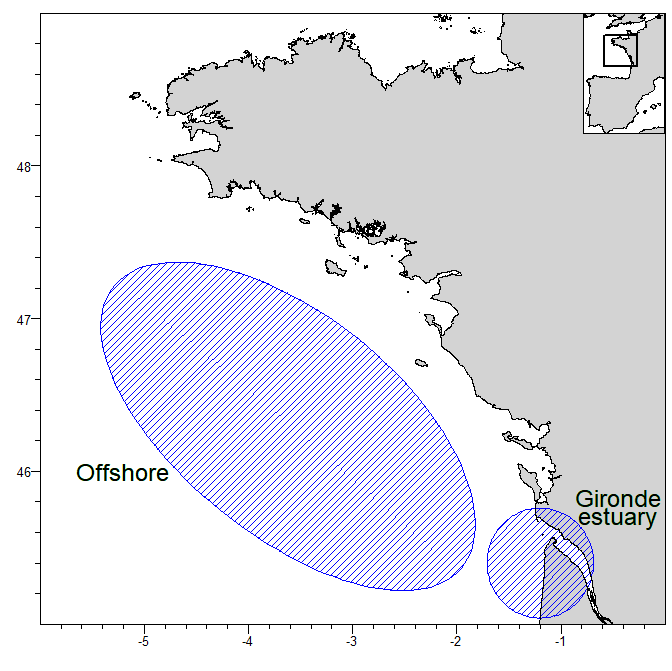
Input data, TMB (C++) and R code for close-kin mark recapture (CKMR) abundance estimation for two subpopulations of thornback ray (Raja clavata) in the central Bay of Biscay (offshore and Gironde estuary, see figure). Samples were taken between 2015 to 2020. Parent-offspring pairs were identified using SNP genotype information. Birth years of individuals were estimated using length information and growth curves by sex.
-
EMODnet Chemistry aims to provide access to marine chemistry datasets and derived data products concerning eutrophication, acidity and contaminants. The importance of the selected substances and other parameters relates to the Marine Strategy Framework Directive (MSFD). This aggregated dataset contains all unrestricted EMODnet Chemistry data on eutrophication and acidity, and covers the Northeast Atlantic Ocean (40W). Data were aggregated and quality controlled by 'IFREMER / IDM / SISMER - Scientific Information Systems for the SEA' in France. ITS-90 water temperature and water body salinity variables have also been included ('as are') to complete the eutrophication and acidity data. If you use these variables for calculations, please refer to SeaDataNet for the quality flags: https://www.seadatanet.org/Products/Aggregated-datasets. Regional datasets concerning eutrophication and acidity are automatically harvested, and the resulting collections are aggregated and quality controlled using ODV Software and following a common methodology for all sea regions ( https://doi.org/10.13120/8xm0-5m67). Parameter names are based on P35 vocabulary, which relates to EMODnet Chemistry aggregated parameter names and is available at: https://vocab.nerc.ac.uk/search_nvs/P35/. When not present in original data, water body nitrate plus nitrite was calculated by summing all nitrate and nitrite parameters. The same procedure was applied for water body dissolved inorganic nitrogen (DIN), which was calculated by summing all nitrate, nitrite, and ammonium parameters. Concentrations per unit mass were converted to a unit volume using a constant density of 1.025 kg/L. The aggregated dataset can also be downloaded as an ODV collection and spreadsheet, which is composed of a metadata header followed by tab separated values. This spreadsheet can be imported to ODV Software for visualisation (more information can be found at: https://www.seadatanet.org/Software/ODV).
-
This dataset provides a World Ocean Atlas of Argo inferred statistics. The primary data are exclusively Argo profiles. The statistics are done using the whole time range covered by the Argo data, starting in July 1997. The atlas is provided with a 0.25° resolution in the horizontal and 63 depths from 0 m to 2,000 m in the vertical. The statistics include means of Conservative Temperature (CT), Absolute Salinity, compensated density, compressiblity factor and vertical isopycnal displacement (VID); standard deviations of CT, VID and the squared Brunt Vaisala frequency; skewness and kurtosis of VID; and Eddy Available Potential Energy (EAPE). The compensated density is the product of the in-situ density times the compressibility factor. It generalizes the virtual density used in Roullet et al. (2014). The compressibility factor is defined so as to remove the dependency with pressure of the in-situ density. The compensated density is used in the computation of the VID and the EAPE.
-
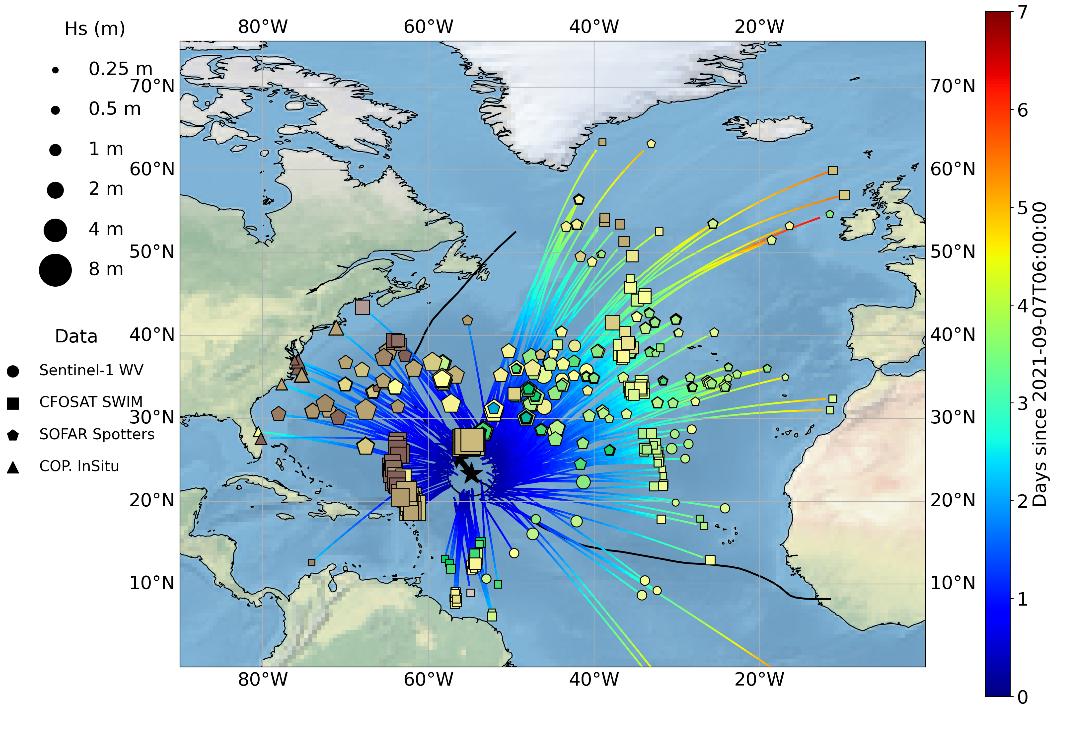
The main objectives of this dataset is to gather the ocean swells measured by different sensors, including satellite and in-situ sources, that were generated by a given tropical cyclone (TC). This dataset aims at providing characteristics of these swells such as their direction, wavelength (or period) and energy but also the date when they left the influence of the tropical cyclone wind to propagate freely. Wave spectra in tropical cyclones vary strongly per quadrant and provide information about the current and past state of the wave field. However, inside TCs, waves measurements including the wave system direction, energy and wavelength are rare and difficult to obtain with in-situ and remote sensing technics. For this dataset, both moored and drifting buoys are considered as long as they provide wave systems measurements. For the satellite contribution, Synthetic Aperture Radar (SAR) and real aperture radar (RAR) instruments can significantly contribute to the TC-generated waves documentation. Indeed, ocean wave spectra can be derived from modulations of the backscatter in SAR and RAR signal. SAR on board European satellite and in particular the SAR series developed since ERS-1 by ESA and now ESA/Copernicus with Sentinel-1 mission (S-1) are good candidates to provide these ocean waves systems characteristics thanks to the dedicated acquisition mode : the so-called Wave Mode. The wave spectrometer SWIM developed by the French space Agency (CNES) and embedded on the Chinese-French Oceanography SATellite (CFOSAT) has been launched more recently with a new measurement concept relying on a RAR and can certainly complement the S-1 data collection. Although the reasons are different, these two systems are limited for measuring waves generation area within the TC vortex where strong rain rates and wind regimes are observed. Far enough from their source, satellite acquisitions are thus expected to be able to observe these ocean swells during more favorable met-ocean conditions for waves retrieval inversion. As a consequence, our analysis is focused on waves originating from TC but that have been able to propagate far from their source. The analysis of swell measurements far from their area of generation to locate the storm source has been firstly applied to data from one single in-situ wave station (wave energy with frequency and direction) collected 2 miles off shore from San Clemente Island, California and extended to a network of several wave stations in the sixties. More recently, the gathering of swell system observed with SAR far from a storm to characterize the waves properties across the ocean has proven to be efficient in the case of extra-tropical storms. Yet, such analysis is not adapted to Tropical Cyclone whose size is much smaller and currently existing wave datasets do not allow for an accurate monitoring of the tropical cyclones swells. This multi-sensor Level-3 tropical cyclone waves dataset intends to fill this gap and opens for an alternate way of estimating tropical cyclone waves properties over all ocean basins and for all tropical cyclones. This dataset was produced in the frame of the ESA funded Marine Atmosphere eXtreme Satellite Synergy (MAXSS) project. The primary objective of the ESA Marine Atmosphere eXtreme Satellite Synergy (MAXSS) project is to provide guidance and innovative methodologies to maximize the synergetic use of available Earth Observation data (satellite, in situ) to improve understanding about the multi-scale dynamical characteristics of extreme air-sea interaction.
-
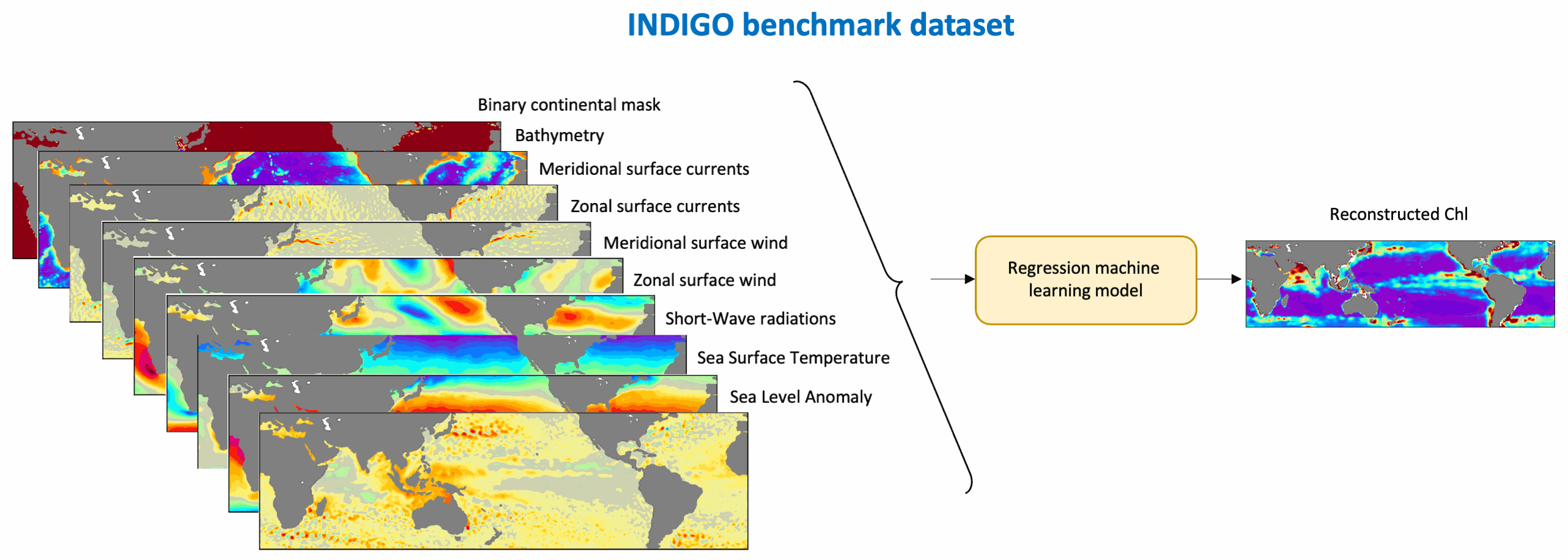
This benchmark dataset contains the physical data used as predictors to reconstruct global chlorophyll-a concentrations (Chl, a proxy of phytoplankton biomass) in Roussillon et al., as well as the reference satellite Chl target fields. The nine physical predictors' data (Short-Wave radiations, Sea Surface Temperature, Sea Level Anomaly, Zonal and meridional surface currents, Zonal and meridional surface wind stress, Bathymetry, Binary continental mask) were extracted from publicly available datasets over [1998-2015] and resampled to the same spatio-temporel resolution as Chl, i.e. monthly on a 1°x1° grid between 50°N and 50°S. Missing values were gap-filled using the heat diffusion equation. Each variable was normalized by substracting its mean from the original values and dividing by its standard deviation over [1998-2015]. This dataset was used to train and validate the Multi-Mode Convolutional Neural network (CNNMM8) introduced in Roussillon et al. ; reconstructed monthly Chl fields over the [2012-2015] test period are also provided here. We hope this benchmark dataset can help to promote the improvements of methods as well as the emergence of new ideas, as building datasets is sometimes more time-consuming than the implementation of machine learning tools themselves. This would also facilitate the quantitative comparison of models performances' on the exact same datasets.
-

'''Short description:''' The product contains a reprocessed multi year version of the daily composite dataset from SEAICE_GLO_SEAICE_L4_NRT_OBSERVATIONS_011_006 covering the Sentinel1 years from autumn 2014 until 1 year before present '''DOI (product) :''' https://doi.org/10.48670/mds-00328
-

'''Short description:''' Altimeter satellite along-track sea surface heights anomalies (SLA) computed with respect to a twenty-year [1993, 2012] mean with a 1Hz (~7km) and 5Hz (~1km) sampling. It serves in near-real time applications. This product is processed by the DUACS multimission altimeter data processing system. It processes data from all altimeter missions available (e.g. Sentinel-6A, Jason-3, Sentinel-3A, Sentinel-3B, Saral/AltiKa, Cryosat-2, HY-2B). The system exploits the most recent datasets available based on the enhanced OGDR/NRT+IGDR/STC production. All the missions are homogenized with respect to a reference mission. Part of the processing is fitted to the European Seas. (see QUID document or http://duacs.cls.fr [http://duacs.cls.fr] pages for processing details). The product gives additional variables (e.g. Mean Dynamic Topography, Dynamic Atmospheric Correction, Ocean Tides, Long Wavelength Errors) that can be used to change the physical content for specific needs (see PUM document for details) '''Associated products''' A time invariant product http://marine.copernicus.eu/services-portfolio/access-to-products/?option=com_csw&view=details&product_id=SEALEVEL_GLO_PHY_NOISE_L4_STATIC_008_033 [http://marine.copernicus.eu/services-portfolio/access-to-products/?option=com_csw&view=details&product_id=SEALEVEL_GLO_PHY_NOISE_L4_STATIC_008_033] describing the noise level of along-track measurements is available. It is associated to the sla_filtered variable. It is a gridded product. One file is provided for the global ocean and those values must be applied for Arctic and Europe products. For Mediterranean and Black seas, one value is given in the QUID document. '''DOI (product) :''' https://doi.org/10.48670/moi-00140
-

In European sea bass like in other animals, the tongue plays a fundamental role in the mechanics of food ingestion. It is composed from the surface in depth of mucosa, submucosa, musculature and fibro cartilaginous skeleton. The tunica mucosa exhibits a stratified epithelium interrupted by numerous teeth differently distributed that erupt more or less completely from the layers below. The European sea bass tongue is composed of canine-like teeth, surrounded by taste buds and numerous fungiform and conical papillae. The tongue beeing directly in contact with external environment, the success of the adaptation of fishes to different environments in the context of global change, depends oamong other on the modifications occurring on the tongue structures. The present study investigates the potential effect of ocean acidification on the lingual transcriptome.
-
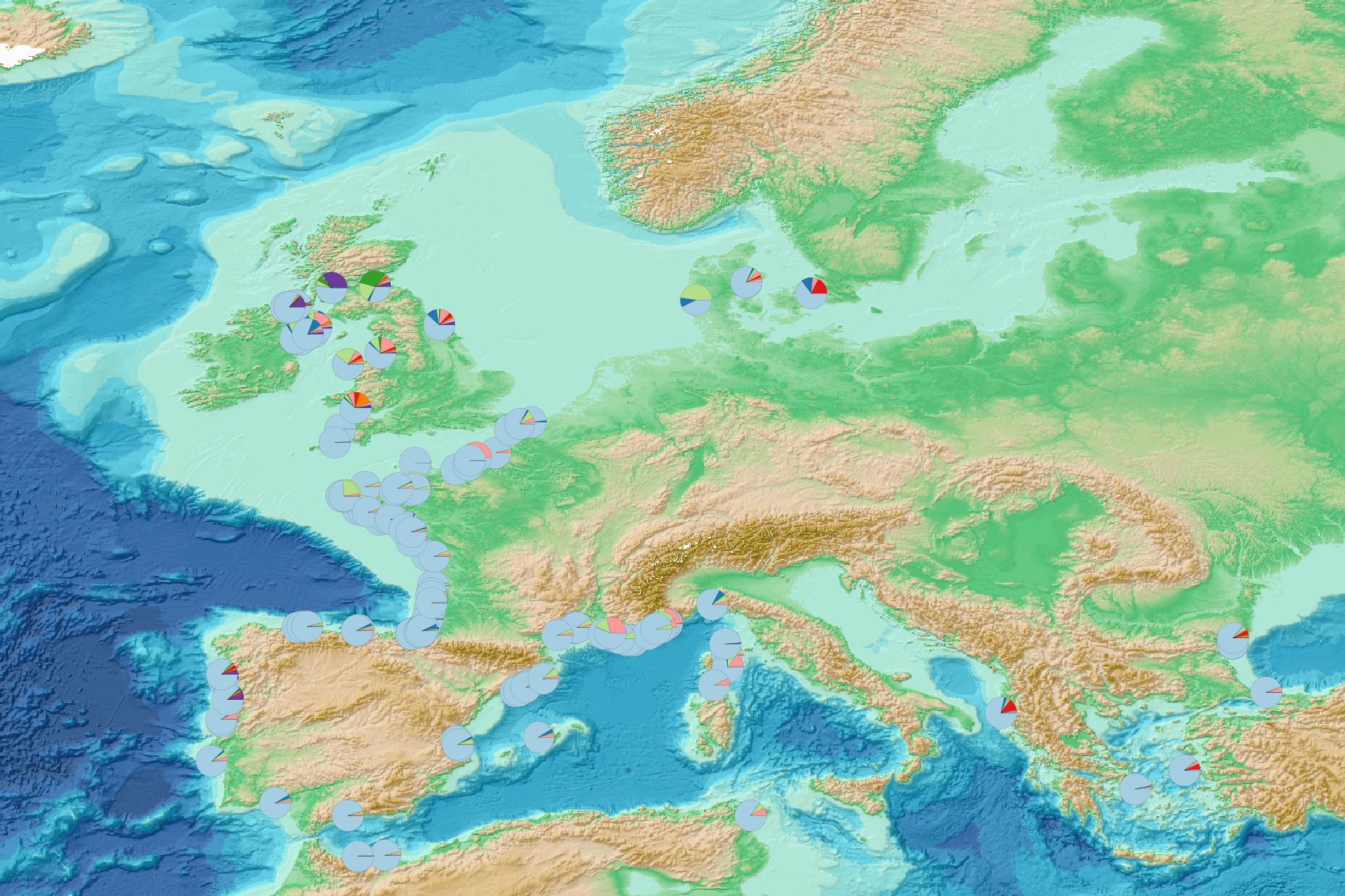
This visualization product displays marine macro-litter (> 2.5cm) material categories percentage per beach per year from non-MSFD monitoring surveys, research & cleaning operations. EMODnet Chemistry included the collection of marine litter in its 3rd phase. Since the beginning of 2018, data of beach litter have been gathered and processed in the EMODnet Chemistry Marine Litter Database (MLDB). The harmonization of all the data has been the most challenging task considering the heterogeneity of the data sources, sampling protocols and reference lists used on a European scale. Preliminary processing were necessary to harmonize all the data: - Exclusion of OSPAR 1000 protocol: in order to follow the approach of OSPAR that it is not including these data anymore in the monitoring; - Selection of surveys from non-MSFD monitoring, cleaning and research operations; - Exclusion of beaches without coordinates; - Exclusion of surveys without associated length; - Some litter types like organic litter, small fragments (paraffin and wax; items > 2.5cm) and pollutants have been removed. The list of selected items is attached to this metadata. This list was created using EU Marine Beach Litter Baselines and EU Threshold Value for Macro Litter on Coastlines from JRC (these two documents are attached to this metadata); - Exclusion of the "feaces" category: it concerns more exactly the items of dog excrements in bags of the OSPAR (item code: 121) and ITA (item code: IT59) reference lists; - Normalization of survey lengths to 100m & 1 survey / year: in some case, the survey length was not 100m, so in order to be able to compare the abundance of litter from different beaches a normalization is applied using this formula: Number of items (normalized by 100 m) = Number of litter per items x (100 / survey length) Then, this normalized number of items is summed to obtain the total normalized number of litter for each survey. To calculate percentages for each material category, formula applied is: Material (%) = (∑number of items (normalized at 100 m) of each material category)*100 / (∑number of items (normalized at 100 m) of all categories) The material categories differ between reference lists (OSPAR, TSG_ML, UNEP, UNEP_MARLIN). In order to apply a common procedure for all the surveys, the material categories have been harmonized. More information is available in the attached documents. Warning: the absence of data on the map doesn't necessarily mean that they don't exist, but that no information has been entered in the Marine Litter Database for this area.
-
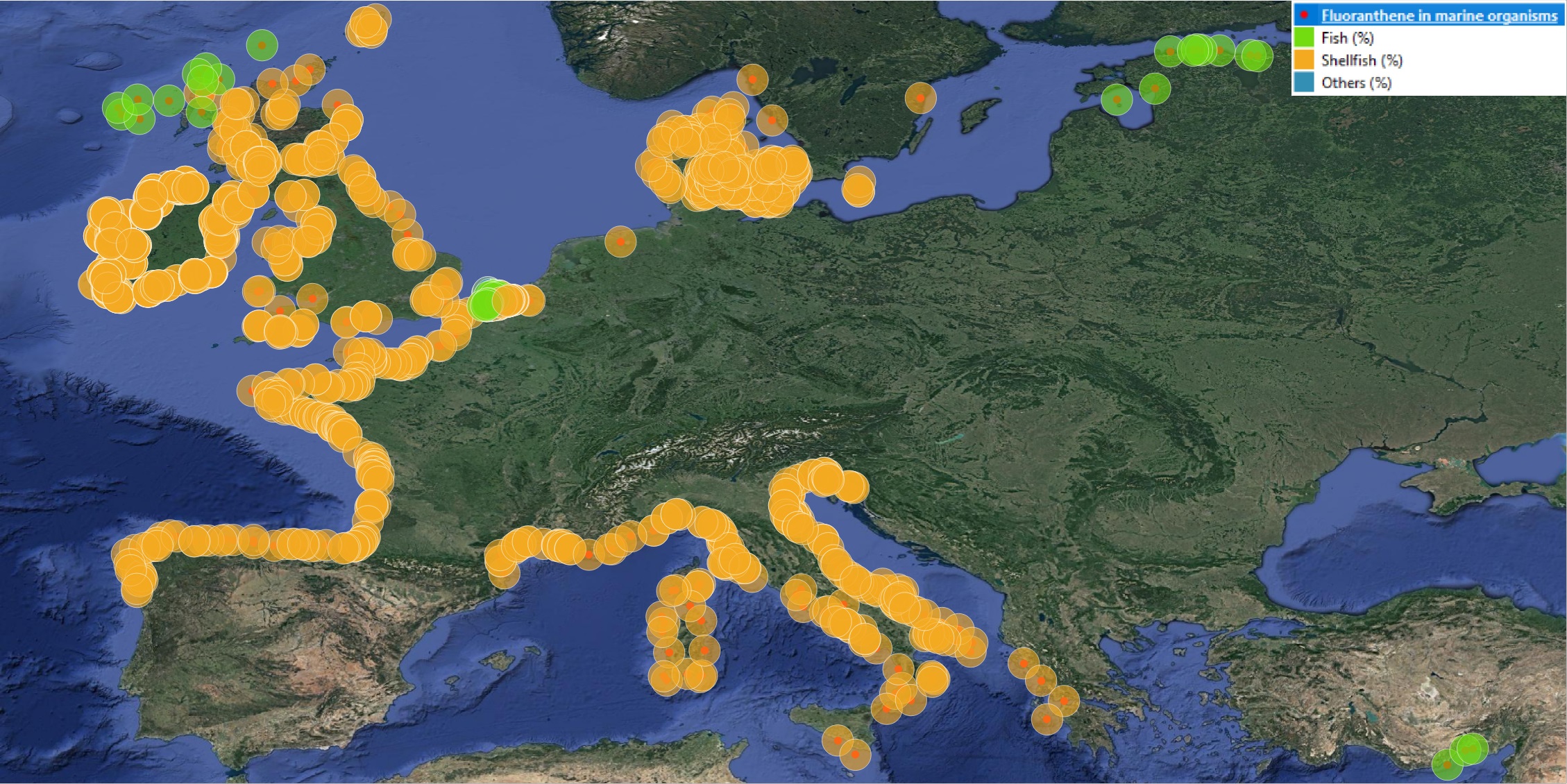
This product displays for Fluoranthene, positions with percentages of all available data values per group of animals that are present in EMODnet regional contaminants aggregated datasets, v2022. The product displays positions for all available years.