 Catalogue PIGMA
Catalogue PIGMA
XLS, XLSX
Type of resources
Available actions
Topics
Keywords
Contact for the resource
Provided by
Years
Formats
-
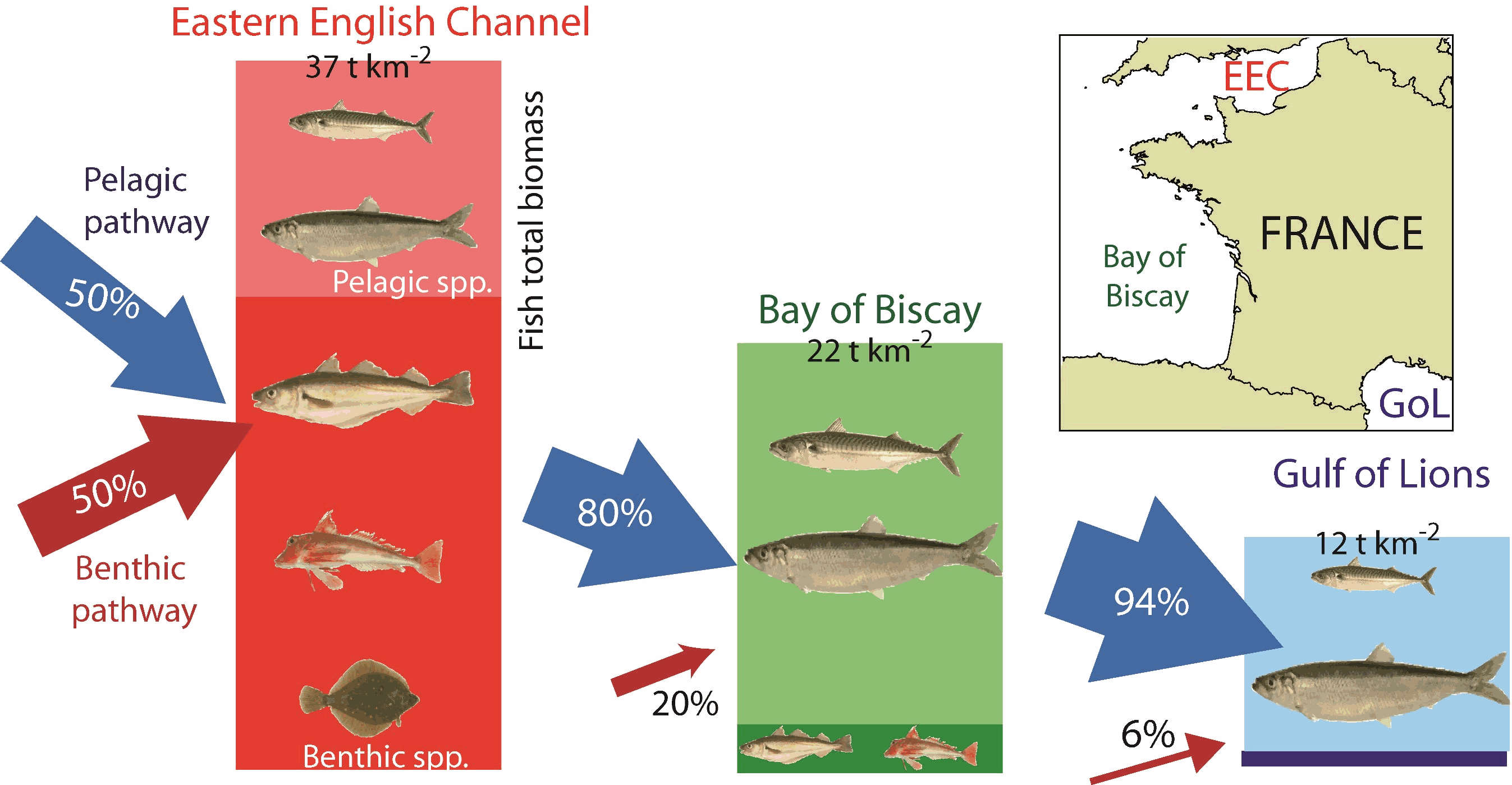
This dataset gathers data used to infer the trophic structure and functioning of fish assemblages in the Eastern English Channel, the Bay of Biscay and the Gulf of Lions : - Biomass data, resulting from accoustic monitoring for pelagic species, or bottom trawling for demersal species, after extrapolation based on stratification scheme - Individual C and N isotopic ratios, length and mass, for all individuals considered - Individual energetic density values
-
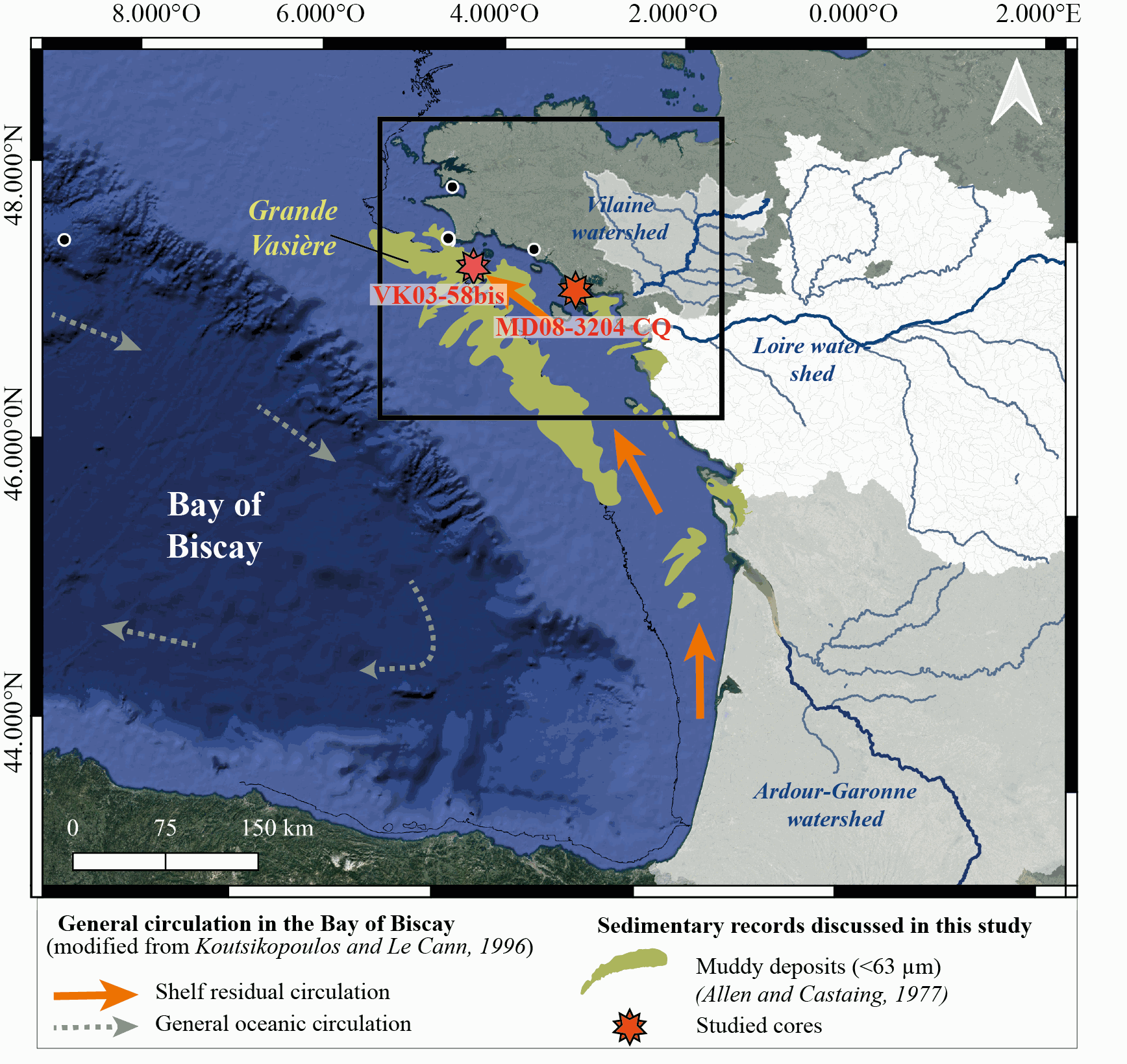
New results acquired in south-Brittany (MD08-3204 CQ core: Bay of Quiberon and VK03-58bis core: south Glénan islands) allow depicting Holocene paleoenvironmental changes from 8.5 ka BP to present through a multi-proxy dataset including sedimentological and palynological data. First, grain-size analyses and AMS-14C dates highlight a common sedimentary history for both study cores. The relative sea level (RSL) slowdown was accompanied by a significant drop of the sedimentation rates between ca. 8.3 and 5.7 ka BP, after being relatively higher at the onset of the Holocene. This interval led to the establishment of a shell-condensed level, identified in core VK03-58bis by the “Turritella layer” and interpreted as a marker for the maximum flooding surface. Palynological data (pollen grains and dinoflagellate cyst assemblages) acquired in core MD08-3204 CQ argue for an amplification of the fluvial influence since 5.7 ka BP; the establishment of the highstand system tract (i.e., mixed marine and fluviatile influences on the platform) then accompanying the slowdown of the RSL rise-rates. On the shelf, the amplification of Anthropogenic Pollen Indicators (API) is then better detected since 4.2 ka BP, not only due to human impact increase but also due to a stronger fluvial influence on the shelf during the Late Holocene. Palynological data, recorded on the 8.5–8.3 ka BP interval along an inshore-offshore gradient, also demonstrate the complexity of the palynological signal such as i) the fluvial influence that promotes some pollinic taxa (i.e., Corylus, Alnus) from proximal areas and ii) the macro-regionalization of palynomorph sources in distal cores. In addition, the comparison of palynological tracers, including API, over the last 7 kyrs, with south-Brittany coastal and mid-shelf sites subjected to northern vs. southern Loire catchment areas, allowed discussing a major hydro-climatic effect on the reconstructed palynological signals. Strengthened subpolar gyre dynamics (SPG), combined with recurrent positive North Atlantic Oscillation (NAO) configurations, appear responsible for increased winter precipitations and fluvial discharges over northern Europe, such as in Brittany. Conversely, weakened SPG intervals, associated with negative NAO-like modes, are characterized by intensified winter fluvial discharges over southern Europe. Interestingly, we record, at an infra-orbital timescale, major peaks of API during periods of strengthened (/weakened) SPG dynamics in sites subjects to Brittany watersheds (/Loire watersheds) inputs.
-

Questions: Invasiveness depends in part on the ability of exotic species to either exclude native dominants or to fill an empty niche. Comparisons of niches and effects of closely related native and invasive species enable the investigation of this topic. Does Spartina anglica invade European salt marshes through competitive exclusion of the native Spartina maritima or due to the occurrence of an empty ecological niche in highly anoxic conditions? Location: The Arcachon Bay (France). Methods: At three intertidal levels, we quantified competitive response and effect abilities of the two species through a cross-transplantation removal experiment. We also compared at three intertidal levels the biomass, root/shoot ratio, productivity and environmental conditions (elevation, salinity, potential redox and soil moisture) of salt marsh communities dominated by the exotic Spartina anglica or the native Spartina maritima. Results: Both established species showed similar biotic resistance to the invasion of the other species, but the exotic showed important intraspecific facilitation for growth. Species had similar niches and total biomass along a gradient of anoxic conditions, but the exotic had a much higher root/shoot ratio and productivity than the native. Owing to its rhizome density, the exotic showed a high ability to increase sediment oxygenation, likely to explain its important intraspecific facilitation. Conclusions: Our results showed that the invasion success of S. anglica cannot be explained by the competitive exclusion of the native or by its ability to fill an empty niche along a gradient of anoxia. Its behaviour as a self-facilitator invasive engineer is very likely to explain its rapid spread in the Bay and biotic resistance to the colonization of other congeneric species when established in dense patches. Additionally, we suggest that physical disturbance in the marsh communities dominated by the native S. maritima may disrupt its biotic resistance against the invasion of S. anglica.
-

Particularly suited to the purpose of measuring the sensitivity of benthic communities to trawling, a trawl disturbance indicator (de Juan and Demestre, 2012, de Juan et al. 2009) was proposed based on benthic species life history traits to evaluate the sensibility of mega- and epifaunal community to fishing pressure known to have a physical impact on the seafloor (such as dredging and bottom trawling). The selected biological traits were chosen as they determine vulnerability to trawling: mobility, fragility, position on substrata, average size and feeding mode that can easily be related to the fragility, recoverability and vulnerability ecological concepts. Life history traits of species have been defined from the BIOTIC database (MARLIN, 2014) and from information given by Le Pape et al. (2007), Brindamour et al. (2009) and Garcia (2010). For missing life history traits, additional information from literature has been considered. The five categories retained are life history functional traits that were selected based on the knowledge of the response of benthic taxa to trawling disturbance (de Juan and Demestre, 2012). They reflect respectively the possibility to avoid direct gear impact, to benefit from trawling for feeding, to escape gear, to get caught by the net and to resist trawling/dredging action, each of these characteristics being either advantageous or sensitive to trawling. Then, to allow quantitative analysis, a score was assigned to each category: from low vulnerability (0) to high vulnerability (3). The five categories scores were then summed for each taxon (the highly vulnerable taxon could reach the maximum score is 15) and this value may be considered as a species index of sensitivity to trawling disturbance. The scores of 812 taxa commonly found in bottom trawl by-catch in the southern North Sea, English Channel and north-western Mediterranean were described.
-

The West Gironde Mud Patch (WGMP) is a 420-km2 mud belt in the Bay of Biscay, located 25 km off the mouth of the Gironde estuary. This clay-silt feature of 4 m in thickness extends between 30 and 75m water depth, surrounded by the sands and gravels that cover the North Aquitaine continental shelf. Interface cores were collected during JERICOBent-1 cruise (October 2016; Deflandre (2016) doi.org/10.17600/16010400) along two cross-shelf transects for a total of 9 sites. Each sediment core was carefully extruded every 0.5 cm from the top core to 4 cm and every 1 cm below until the core bottom. The sediment layers were used to determine dry bulk density, grain size and selected radioisotope activities (210Pb, 226Ra, 137Cs, 228Th, K).
-
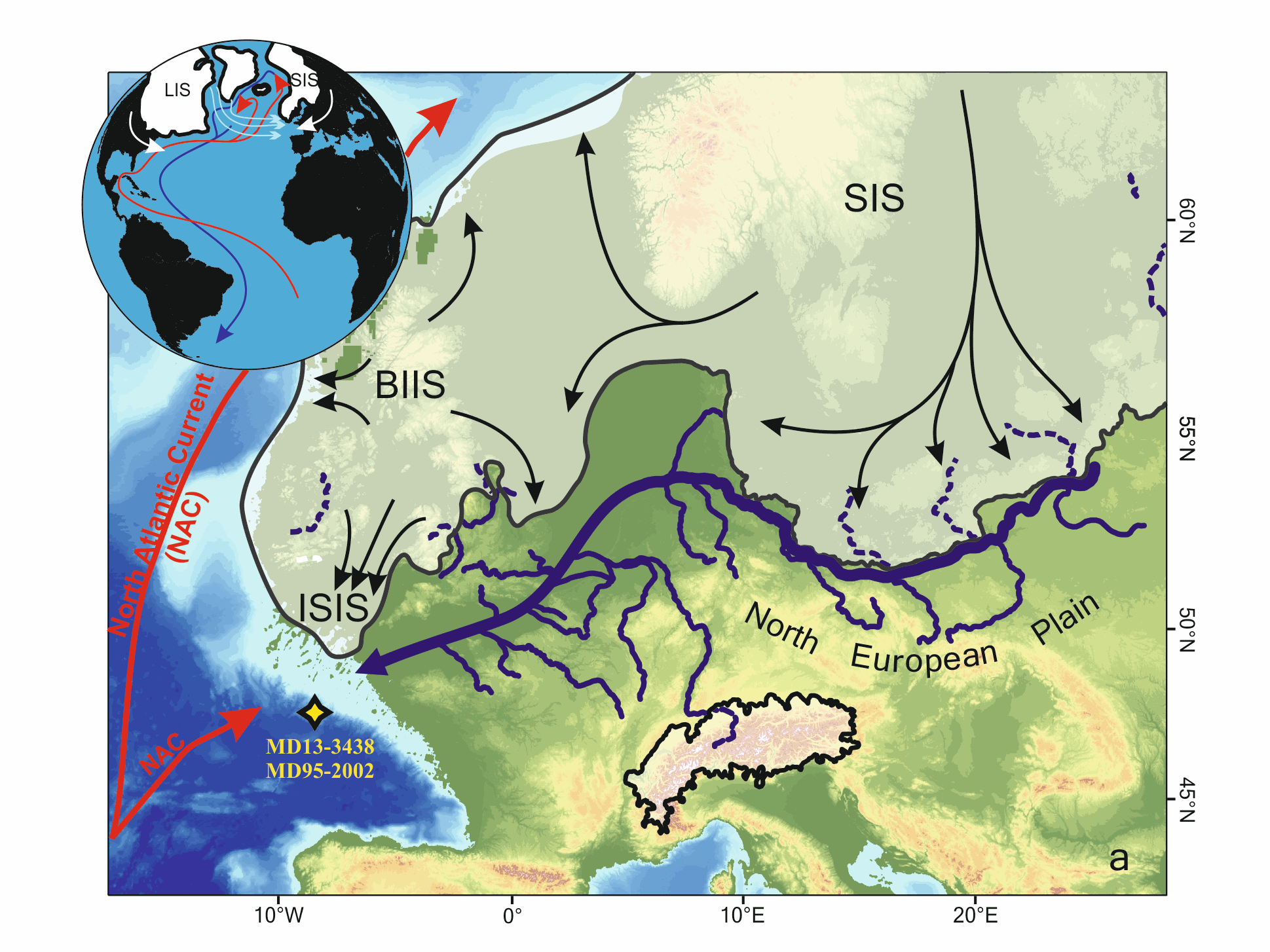
Marine microfossils (dinoflagellate cysts and planktonic foraminifera) and geochemical (XRF-Ti/Ca)-based climatic records from a core located off the Fleuve Manche (FM) paleo-mouth (MD13-3438) have revealed that sustained warm summer sea surface temperatures (SSTs) during sub-millennial climate changes within HS1 (~18–14.7 ka) may have played a key role in the FM regime related to the European Ice Sheet (EIS) melting rate. In this study, we have analyzed the MD13-3438 pollen content over the HS1 at a mean resolution of ~50 years to test whether vegetation-based air temperatures were coupled to SSTs face to this rapid climate variability. First, our results highlight two major phases of pollen sources at site MD13-3438, preventing the pollen record to be interpreted as a continuous record of the evolution of vegetation and climate occupying a single watershed across HS1. The first phase, i.e. the HS1-a interval (~18–16.8 ka), is marked by strong occurrences of boreal pollen taxa (especially Picea-Abies). Considering their spatial distribution and the coalescence of the British and Scandinavian ice sheets into the North Sea during the Last Glacial Maximum, these taxa probably originated from the North European Plain, i.e., eastern FM tributaries (east of the Rhine River), where cool-humid conditions generally prevailed. Then, the second phase, i.e. the HS1-b interval (~16.8–14.7 ka BP), is characterized by a deceleration of the EIS retreat and the drop of boreal pollen values at site MD13-3438 further signing a less influence of the upstream FM drainage system and thus a better characterization of pollen sources related with western FM tributaries. Superimposed to these two HS1 main phases, pollen fluctuations are concomitant with sub-millennial variability in the EIS deglaciation intensity. During the early HS1 (HS1-a), we discussed two short-term increases in the ratio between deciduous trees (Quercus-Corylus-Alnus) and herbaceous plants (Plantago-Amaranthaceae-Artemisia). These events were coeval with phases of increasing FM meltwater runoff and SST seasonality (i.e., dinocyst-based summer SST amplification). We associated these events with lower contribution of the upstream FM catchment as well as, possibly, atmospheric warming and regional sea-level positive oscillations. The HS1-b is composed of three main phases that appear more influenced by the downstream FM drainage system. HS1-b1 (16.8–16.3 ka BP) corresponds to the driest and coldest conditions west of the Rhine River. HS1-b2 (16.3–15.6 ka BP) is coeval with large arrivals of iceberg from the Hudson strait in the Bay of Biscay and thus likely to a major sea-level positive oscillation associated with a phase of FM valley reworking. HS1-b3 (15.6–14.7 ka BP) corresponds to persistent arid conditions that preceded the subsequent more humid conditions recorded from 14.7 ka BP at the start of the Bölling-Alleröd.
-

The ARCHYD dataset, which have been collected since 1988, represents the longest long-term hydrologic data sets in Arcachon Bay. The objectives of this monitoring programme are to assess the influence of oceanic and continental inputs on the water quality of the bay and their implications on biological processes. It also aims to estimate the effectiveness of management policies in the bay by providing information on trends and/or shifts in pressure, state, and impact variables. Sampling is carried on stations spread across the entire bay, but since 1988, the number and location of stations have changed slightly to better take into account the gradient of ocean and continental inputs. In 2005, the ARCHYD network was reduced to 8 stations that are still sampled by Ifremer to date. All the stations are sampled at a weekly frequency, at midday, alternately around the low spring tide and the high neap tide. Data are complementary to REPHY dataset. Physico-chemical measures include temperature, salinity, turbidity, suspended matters (organic, mineral), dissolved oxygen and dissolved inorganic nutrients (ammonium, nitrite+nitrate, phosphate, silicate). Biological measures include pigment proxies of phytoplankton biomass and state (chlorophyll a and phaeopigment).
-
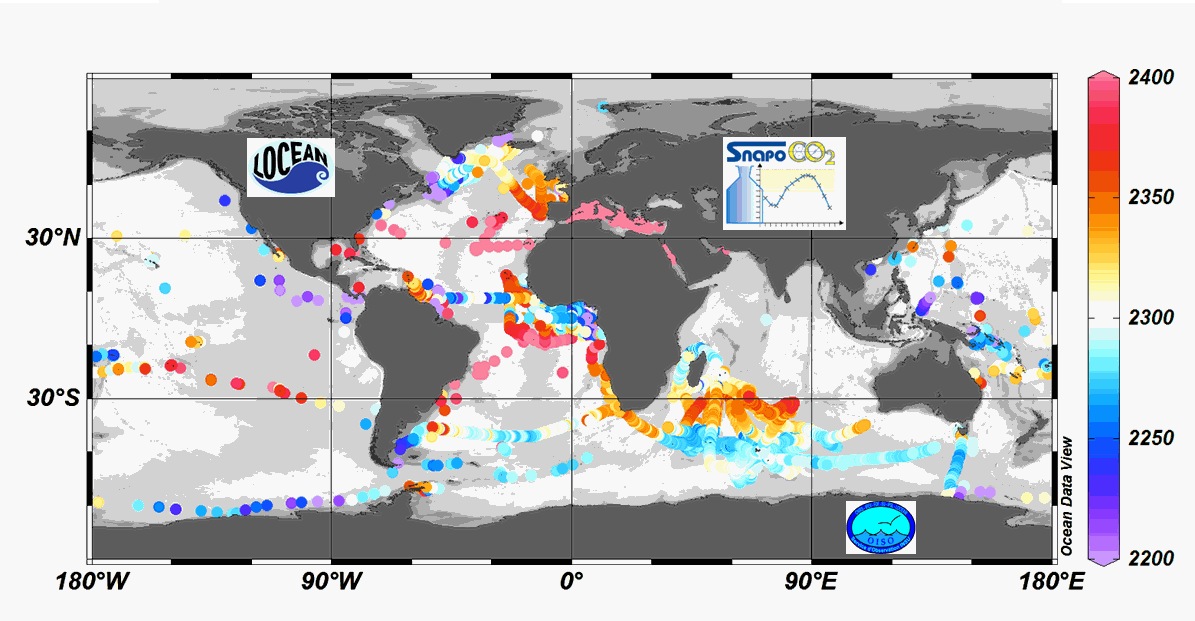
Since 2004, the Service facility SNAPO-CO2 (Service National d’Analyse des Paramètres Océaniques du CO2) housed by the LOCEAN laboratory (Paris, France) has been in charge for the analysis of Total Alkalinity (AT) and Total dissolved inorganic carbon (CT) of seawater samples on a series of cruises or ships of opportunity conducted in different regions in the frame of French projects. More than 44000 observations are synthetized in this work. Sampling was performed either from CTD-Rosette casts (Niskin bottles) or collected from the ship’s seawater supply (intake at about 5m depth). After completion of each cruise, discrete samples were returned back at LOCEAN laboratory and stored in a dark room at 4 °C before analysis generally within 2-3 months after sampling (sometimes within a week). AT and CT were analyzed simultaneously by potentiometric titration using a closed cell (Edmond, 1970). Certified Reference Materials (CRMs) provided by Pr. A. Dickson (Scripps Institution of Oceanography, San Diego, USA) were used to calibrate the measurements. The same instrumentation was used for underway measurements during OISO cruises (https://doi.org/10.18142/228) and OISO AT-CT data for 1998-2018 in the South Indian Ocean added in this synthesis. The synthesis is organized in two files (one for Global ocean and the Coastal Zones, one for the Mediterranean Sea) with the same format: Cruise name, Ship name, day, month, year, hour, minute, second, latitude, longitude, depth, AT (µmol/kg), Flag-AT, CT (µmol/kg), Flag-CT, Temperature (°C), Flag-Temp, Salinity (PSU), Flag-Salinity, nsample/cruise, nsample on file, sampling method.
-
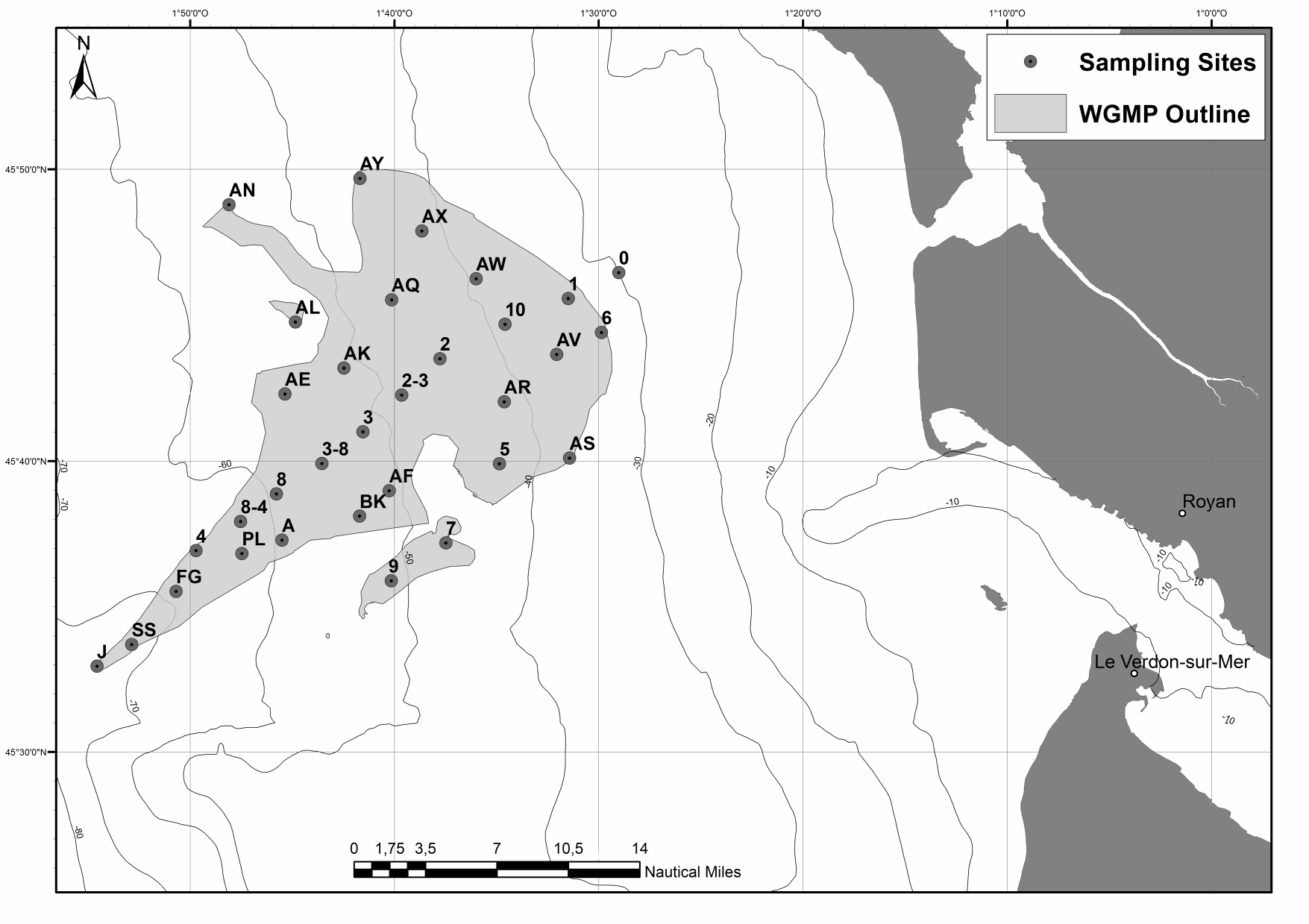
The spatial distributions of (1) surface sediment characteristics (D0.5, Sediment Surface Area (SSA), Particulate Organic Carbon (POC), Chlorophyll-a (Chl-a), Phaeophytin-a (Phaeo-a), Total and Enzymatically Hydrolyzable Amino Acids (THAA, EHAA), δ13C) and (2) sediment profile image (apparent Redox Potential Discontinuity (aRPD), numbers and depths of biological traces) characteristics were quantified based on the sampling of 32 stations located within the West Gironde Mud Patch (Bay of Biscay, NE Atlantic) in view of (1) assessing the spatial structuration of a temperate river-dominated ocean margin located in a high-energy area, (2) disentangling the impacts of hydrodynamics and bottom trawling on this structuration, and (3) comparing the West Gironde Mud Patch with the Rhône River Prodelta (located in a low-energy area). Results support the subdivision of the West Gironde Mud Patch in a proximal and a distal part and show (1) the existence of depth gradients in surface sedimentary organics characteristics and bioturbation within the distal part; (2) no evidence for a significant effect of bottom trawling, as opposed to Bottom Shear Stress, on the West Gironde Mud Patch spatial structuration; and (3) major discrepancies between spatial structuration in the West Gironde Mud Patch and the Rhône River Prodelta, which were attributed to differences in tidal regimes, sedimentation processes, and local hydrodynamics, which is in agreement with current river-dominated ocean margin typologies.
-

Good Environmental Status assessment (GES) for descriptor 8 (contaminants, D8) of the Marine Strategy Framework Directive (MSFD) is reached when concentrations of contaminants are at levels not giving rise to pollution effects. It is described by 4 criteria among which the first one focus on the concentration of the contaminants in the environment (criteria 1 of the D8, D8C1). The environmental status for D8 in France includes assessment of contaminant concentrations in sediment, bivalves, fish, birds, mammals to cover the French marine area the continental shelf from the coast line). The 8 tables below present the assessment of the chemical contamination in sediment and bivalves on the coastal area of the 4 French marine subregions for D8 as part of the 2024 GES assessment. These tables report the status and temporal trends of each station x matrice x substance triplet in each of the 4 French marine subregions. Explanation on how to read the cells is given in the “read file”. The environmental assessment for D8 in France can be found in Mauffret al., 2023 (DOI:10.13155/97214). It includes 17 national indicator assessments, 4 OSPAR indicators and integrated assessment in selected assessment units at the level of the criteria 1 and 2.