 Catalogue PIGMA
Catalogue PIGMA
Analysis data
Type of resources
Available actions
Topics
Keywords
Contact for the resource
Provided by
Years
Formats
Representation types
-

Comparison between 3 kits, Roche, Twist Biosciences and Illumina on the ability to enrich environmental samples to viral sequences
-

WGS of SARS-CoV-2 by Oxford Nanopore Technology from raw wastewater samples collected in France, 2020-2021
-

Vibrio bacteria sampled from juvenile oysters and seawater collected in Thau Lagoon (Languedoc-Roussillon, France) in October 2015 during a mortality event were genotyped using hsp60, rctB, topA and mreB protein-coding genes
-

160 whole genomes sequences obtained from 160 individual fish samples representing about 100 different species present in Gulf of Lion, and bay of Biscay.
-

This study gathers multi-year environmental sequencing datasets generated within the French ROME pilot observatory network. It includes eDNA metabarcoding and RNA-based analyses from water samples, oyster tissues, and viral fractions collected across four French estuarine ecosystems between 2020 and 2023, supporting integrated monitoring of coastal microbiomes and microbial hazards.
-

DNA sequencing of Crassostrea gigas Pacific oyster spat infected in the wild with OsHV-1 virus in 4 French oyster basins (Marennes Oleron Bay, Arcachon bay, Rade de Brest and Thau lagoon).
-

Raw reads for the assembly of Gambusia holbrooki genome.
-

This dataset consists of metatranscriptomic sequencing reads corresponding to coastal micro-eukaryote communities sampled in Western Europe in 2018 and 2019.
-
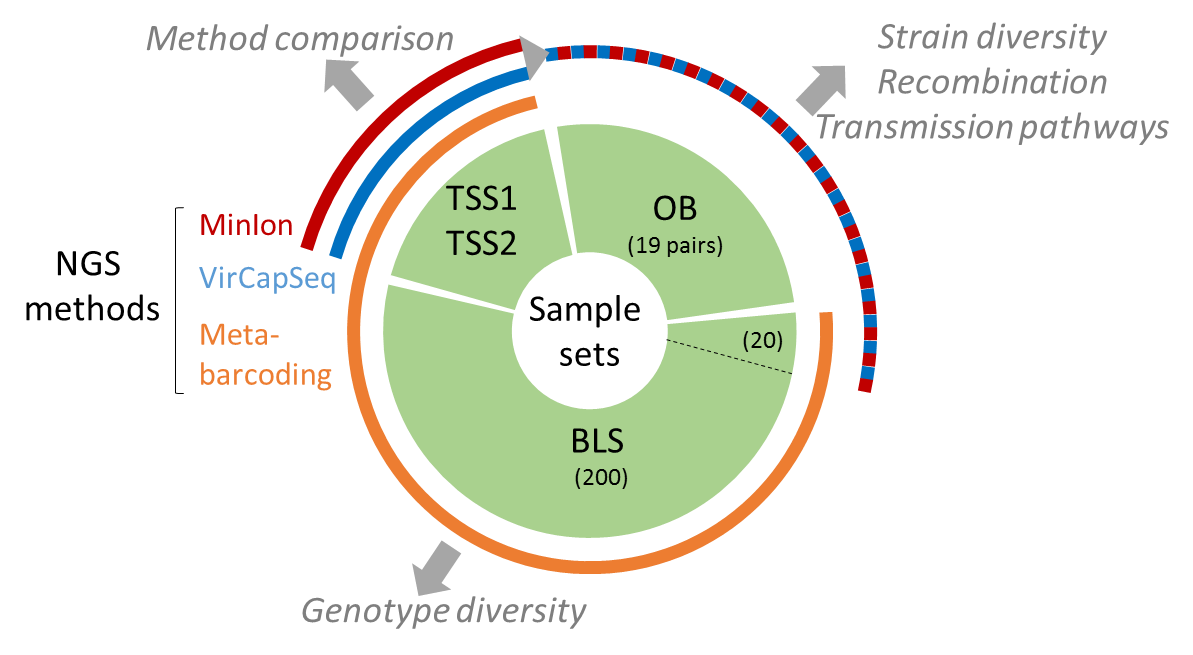
The present data set concerne metabarcoding raw reads produced using 4 different PCR targeting polymerase or capside coding region of the genoyupe I and II of norovirus. Test samples of norovirus with serial dilutions in pure water and after a bio-accumulation in oysters. Sequencing was made after VirCapSeq-VERT approach.
-

This study aims to compare different metabarcoding sequences of commercially fished shrimps collected by tree counties on the North Brazil Shelf Large Marine Ecosystem