 Catalogue PIGMA
Catalogue PIGMA
2022
Type of resources
Available actions
Topics
Keywords
Contact for the resource
Provided by
Years
Formats
Representation types
Update frequencies
status
Service types
Scale
Resolution
-
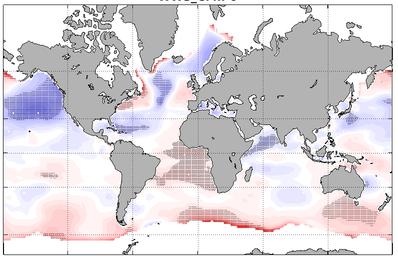
Global wave hindcast (1961-2020) at 1° resolution using CMIP6 wind and sea-ice forcings for ALL (historical), GHG (historical greenhouse-gas-only), AER (historical Anthropogenic-aerosol-only), NAT (historical natural only) scenario.
-
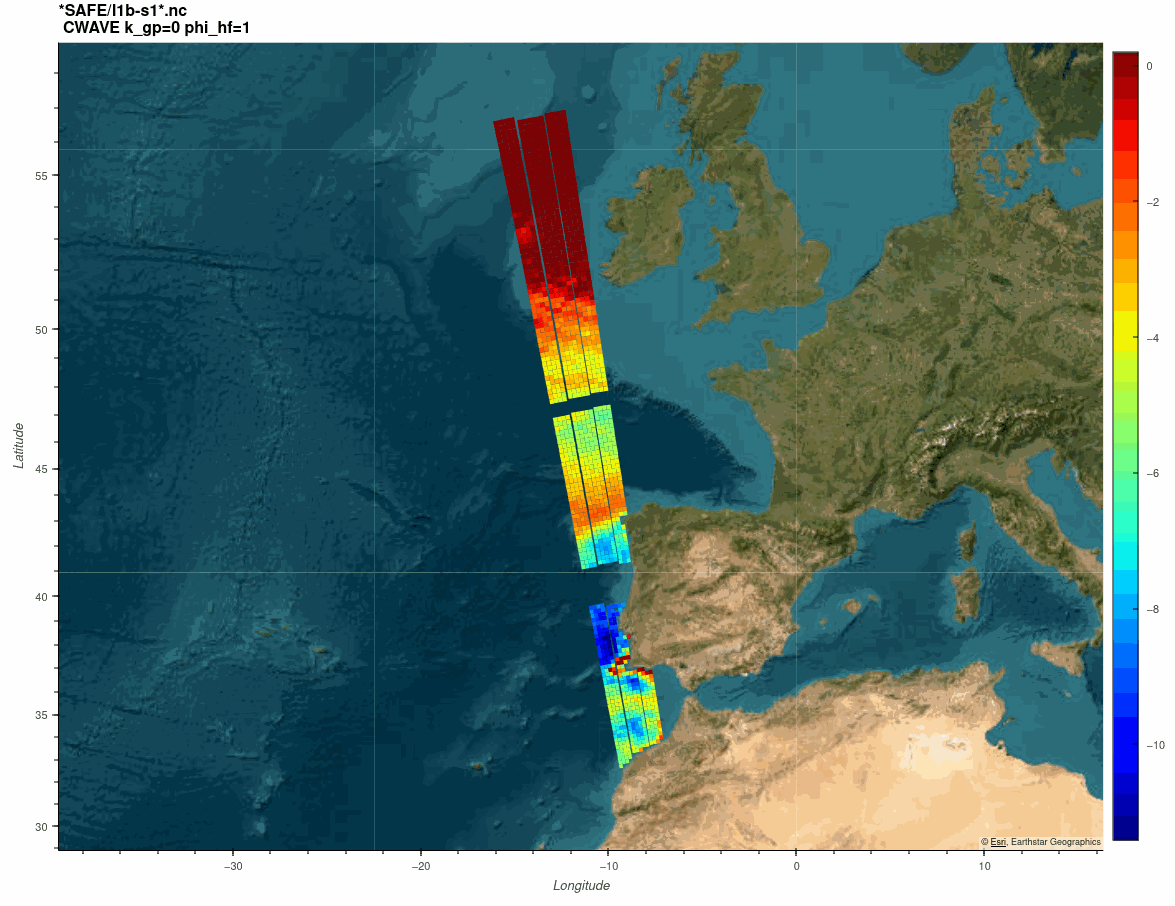
The SARWAVE project is developing a new sea state processor from SAR images to be applied over open ocean, sea ice, and coastal areas, and exploring potential synergy with other microwave and optical EO products.
-
French benthic invertebrates composition and abundance taxa data are collected during monitoring surveys on the English Channel / Bay of Biscay coasts and Mediterranean coast (Quadrige program code : REBENT_FAU, RSL_FAU). Protocols are implemented in the Water Framework Directive. Data are transmitted in a Seadatanet format (CDI + ODV) to EMODnet Biology european database. 498 ODV files have been generated from period 01/01/2003 to 31/12/2021.
-
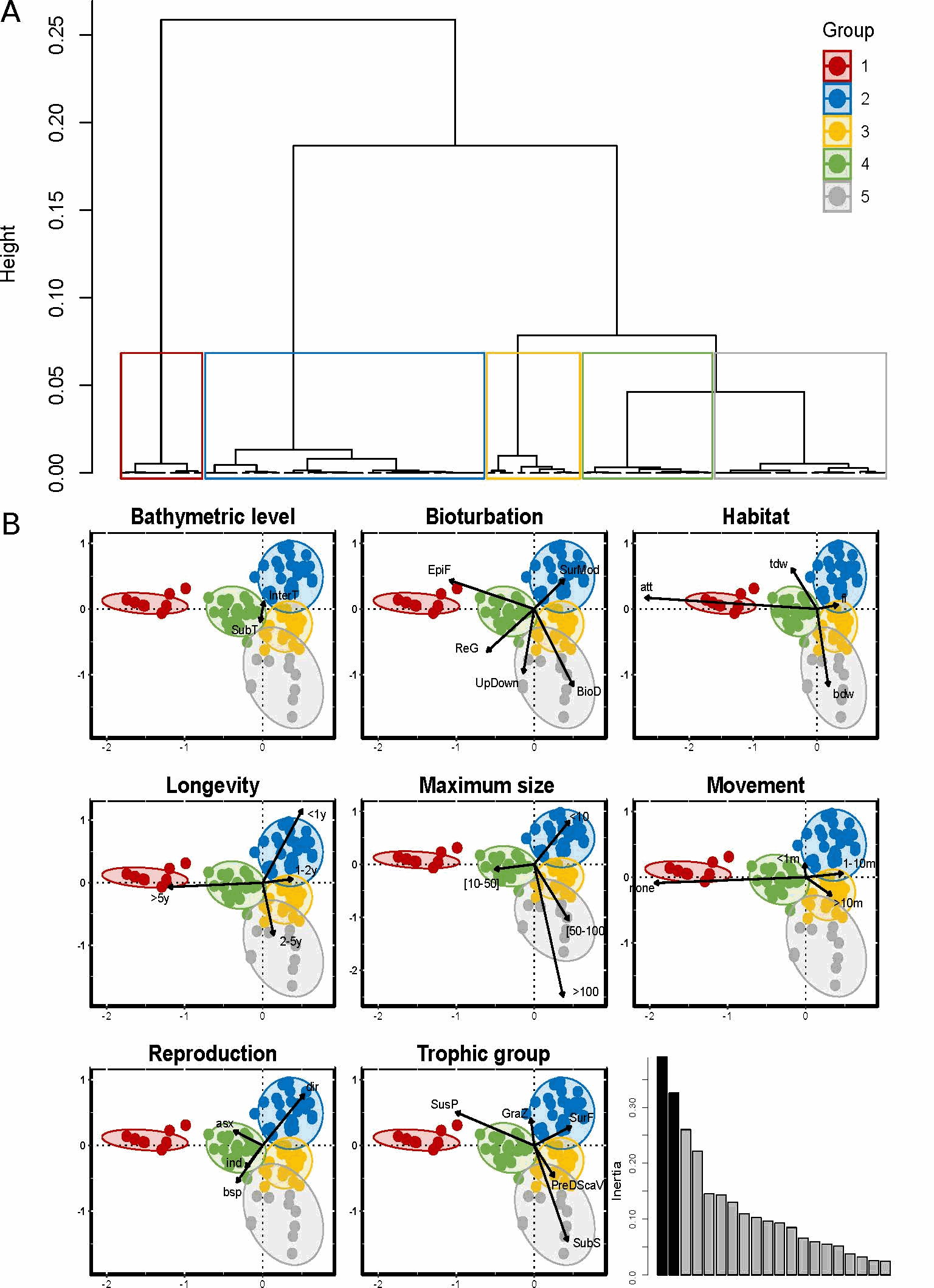
Reef-building species are recognized as having an important ecological role and as generally enhancing the diversity of benthic organisms in marine habitats. However, although these ecosystem engineers have a facilitating role for some species, they may exclude or compete with others. The honeycomb worm Sabellaria alveolata (Linnaeus, 1767) is an important foundation species, commonly found from northwest Ireland to northern Mauritania (Curd et al., 2020), whose reef structures increase the physical complexity of the marine benthos, supporting high levels of biodiversity. Local patterns and regional differences in taxonomic and functional diversity were examined in honeycomb worm reefs from ten sites along the northeastern Atlantic to explore variation in diversity across biogeographic regions and the potential effects of environmental drivers. To characterize the functional diversity at each site, a biological trait analysis (BTA) was conducted (Statzner et al., 1994). Here we present the functional trait database used for the benthic macrofauna found to live in association with honeycomb worm reefs. Eight biological traits (divided into 32 modalities) were selected (Table 1), providing information linked to the ecological functions performed by the associated macrofauna. The selected traits provide information on: (i) resource use and availability (by the trophic group of species, e.g. Thrush et al. 2006); (ii) secondary production and the amount of energy and organic matter (OM) produced based on the life cycle of the organisms (including longevity, maximum size and mode of reproduction, e.g. (Cusson and Bourget, 2005; Thrush et al., 2006) and; (iii) the behavior of the species in general [i.e. how these species occupy the environment and contribute to biogeochemical fluxes through habitat, movement, and bioturbation activity at different bathymetric levels, e.g. (Solan et al., 2004; Thrush et al., 2006; Queirós et al., 2013). Species were scored for each trait modality based on their affinity using a fuzzy coding approach (Chevenet et al., 1994), where multiple modalities can be attributed to a species if appropriate, and allowed for the incorporation of intraspecific variability in trait expression. The information concerning polychaetes was derived primarily from Fauchald et al (1979) and Jumars et al (2015). Information on other taxonomic groups was obtained either from databases of biological traits (www.marlin.ac.uk/biotic) or publications (Naylor, 1972; King, 1974; Caine, 1977; Lincoln, 1979; Holdich and Jones, 1983; Smaldon et al., 1993; Ingle, 1996; San Martín, 2003; Southward, 2008; Gil, 2011; Leblanc et al., 2011; Rumbold et al., 2012; San Martín and Worsfold, 2015; Jones et al., 2018). Map indicating the locations of the 10 study sites in the UK, France and Portugal within the four biogeographic provinces defined by Dinter (2001). (All sites were sampled in 8 different stations, except for UK4 where 5 stations were sampled).
-
This dataset contains the pictures used for morphometric measurements, as well as the elemental compositon and production rates data, of planktonic Rhizaria. Specimens were collected in the bay of Villefranche-sur-Mer in May 2019 and during the P2107 cruise in the California Current in July-August 2021. Analyses of the data can be found at https://github.com/MnnLgt/Elemental_composition_Rhizaria.
-
The ESA Sea State Climate Change Initiative (CCI) project has produced global multi-sensor time-series of along-track satellite synthetic aperture radar (SAR) integrated sea state parameters (ISSP) data from ENVISAT (referred to as SAR Wave Mode onboard ENVISAT Level 2P (L2P) ISSP data) with a particular focus for use in climate studies. This dataset contains the ENVISAT Remote Sensing Integrated Sea State Parameter product (version 1.1), which forms part of the ESA Sea State CCI version 3.0 release. This product provides along-track significant wave height (SWH) measurements at 5km resolution every 100km, processed using the Li et al., 2020 empirical model, separated per satellite and pass, including all measurements with flags and uncertainty estimates. These are expert products with rich content and no data loss. The SAR Wave Mode data used in the Sea State CCI SAR WV onboard ENVISAT Level 2P (L2P) ISSP v3 dataset come from the ENVISAT satellite mission spanning from 2002 to 2012.
-
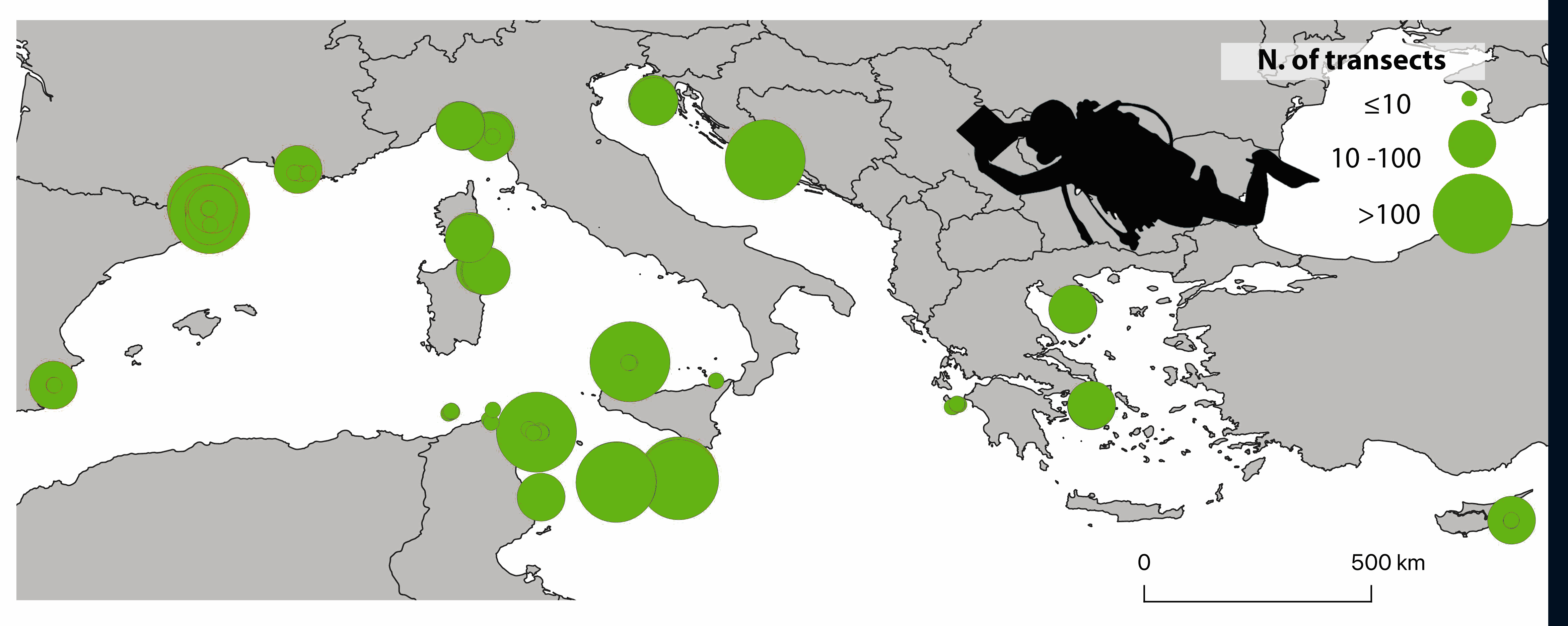
The ClimateFish database collates abundance data of 15 fish species proposed as candidate indicators of climate change in the Mediterranean Sea. An initial group of eight Mediterranean indigenous species (Epinephelus marginatus, Thalassoma pavo, Sparisoma cretense, Coris julis, Sarpa salpa, Serranus scriba, Serranus cabrilla and Caranx crysos) with wide distribution, responsiveness to temperature conditions and easy identification were selected by a network of Mediterranean scientists joined under the CIESM programme ‘Tropical Signals’ (https://www.ciesm.org/marine/programs/tropicalization.htm; Azzurro et al. 2010). Soon after, and thanks to the discussion with other expert groups and projects, C. crysos was no longer considered, and Lessepsian fishes (Red Sea species entering the Mediterranean through the Suez Canal) were included, namely: Fistularia commersonii, Siganus luridus, Siganus rivulatus, Pterois miles, Stephanolopis diaspros, Parupeneus forskali, Pempheris rhomboidea and Torquigener flavimaculosus. Considering the trend of increase of these species in the Mediterranean Sea (Golani et al. 2021) and their projected distribution according to climate change scenarios (D’Amen and Azzurro, 2020), more data on these tropical invaders are expected to come in the future implementation of the study. Data were collected according to a simplified visual census methodology (Garrabou et al. 2019) along standard transects of five minutes performed at a constant speed of 10m/min, corresponding approximately to an area of 50x5m. Four different depth layers were surveyed: 0-3m, 5-10 m, 11-20 m, 21-30 m. So far, the ClimateFish database includes fish counts collected along 3142 transects carried out in seven Mediterranean countries between 2009 and 2021, for a total number of 101'771 observed individuals belonging to the 15 fish species. Data were collected by a large team of researchers which joined in a common monitoring strategy supported by different international projects, which are acknowledged below. This database, when associated with climate data, offers new opportunities to investigate spatio-temporal effects of climate change in the Mediterranean Sea and test the effectiveness of each species as a possible climate change indicator. Contacts: ernesto.azzurro(at)cnr.it References: Azzurro E., Maynou F., Moschella P. (2010). A simplified visual census methodology to detect variability trends of coastal mediterranean fishes under climate change scenarios. Rapp. Comm. int. Mer Médit., 39. D’Amen, M. and Azzurro, E. (2020). Lessepsian fish invasion in Mediterranean marine protected areas: a risk assessment under climate change scenarios. ICES Journal of Marine Science, 77(1), pp.388-397. Garrabou, J., Bensoussan, N., Azzurro, E. (2019). Monitoring climate-related responses in Mediterranean marine protected areas and beyond: five standard protocols. Golani D., Azzurro E., Dulčić J., Massutí E., Orsi-Relini L. (2021). Atlas of Exotic Fishes in the Mediterranean Sea. 2nd edition [F. Briand, Ed.] 365 pages. CIESM Publishers, Paris, Monaco. ISBN number 978-92-990003-5-9
-
This dataset provides surface Stokes drift as retrieved from the wave energy spectrum computed by the spectral wave model WAVEWATCH-III (r), under NOAA license, discretized in wave numbers and directions and the water depth at each location. It is estimated at the sea surface and expressed in m.s-1. WAVEWATCH-III (r) model solves the random phase spectral action density balance equation for wavenumber-direction spectra. Please refer to the WAVEWATCH-III User Manual for fully detailed description of the wave model equations and numerical approaches. The data are available through HTTP and FTP; access to the data is free and open. In order to be informed about changes and to help us keep track of data usage, we encourage users to register at: https://forms.ifremer.fr/lops-siam/access-to-esa-world-ocean-circulation-project-data/ This dataset was generated by Ifremer / LOPS and is distributed by Ifremer / CERSAT in the frame of the World Ocean Circulation (WOC) project funded by the European Space Agency (ESA).
-

The French Atlantic coast hosts numerous macrotidal and turbid estuaries that flow into the Bay of Biscay that are natural corridors for migratory fishes. The two best known are those of the Gironde and the Loire. However, there are also a dozen estuaries set geographically among them, of a smaller scale. The physico-chemical quality of estuarine waters is a necessary support element for biological life and determines the distribution of species, on which many ecosystem services (e.g. professional or recreational fishing) depend. With rising temperatures and water levels, declining precipitation and population growth projected for the New Aquitaine region by 2030, the question of how the quality and ecological status of estuarine waters will evolve becomes increasingly critical. The MAGEST (Mesures Automatisées pour l’observation et la Gestion des ESTuaires nord aquitains) high-frequency monitoring of key physico-chemical parameters was first developed in the Gironde estuary in 2004 ; the Seudre and Charente estuaries were instrumented late 2020. First based on real-time automated systems, MAGEST is now equipped by autonomous multiparameter sensors. Depending of the stations, an optode is also deployed to secure dissolved oxygen measurement. By the end of 2020, MAGEST had 12 instrumented sites. Portets is a measuring station located in the upper Gironde estuary (Garonne subestuary, about 20 km upstream of the Bordeaux metropolis.
-
The GEBCO_2022 Grid is a global continuous terrain model for ocean and land with a spatial resolution of 15 arc seconds. In regions outside of the Arctic Ocean area, the grid uses as a base Version 2.4 of the SRTM15_plus data set (Tozer, B. et al, 2019). This data set is a fusion of land topography with measured and estimated seafloor topography. Included on top of this base grid are gridded bathymetric data sets developed by the four Regional Centers of The Nippon Foundation-GEBCO Seabed 2030 Project. The GEBCO_2022 Grid represents all data within the 2022 compilation. The compilation of the GEBCO_2022 Grid was carried out at the Seabed 2030 Global Center, hosted at the National Oceanography Centre, UK, with the aim of producing a seamless global terrain model. Outside of Polar regions, the Regional Centers provide their data sets as sparse grids i.e. only grid cells that contain data are populated. These data sets were included on to the base using a remove-restore blending procedure. This is a two-stage process of computing the difference between the new data and the base grid and then gridding the difference and adding the difference back to the existing base grid. The aim is to achieve a smooth transition between the new and base data sets with the minimum of perturbation of the existing base data set. The data sets supplied in the form of complete grids (primarily areas north of 60N and south of 50S) were included using feather blending techniques from GlobalMapper software. The GEBCO_2022 Grid has been developed through the Nippon Foundation-GEBCO Seabed 2030 Project. This is a collaborative project between the Nippon Foundation of Japan and the General Bathymetric Chart of the Oceans (GEBCO). It aims to bring together all available bathymetric data to produce the definitive map of the world ocean floor by 2030 and make it available to all. Funded by the Nippon Foundation, the four Seabed 2030 Regional Centers include the Southern Ocean - hosted at the Alfred Wegener Institute, Germany; South and West Pacific Ocean - hosted at the National Institute of Water and Atmospheric Research, New Zealand; Atlantic and Indian Oceans - hosted at the Lamont-Doherty Earth Observatory, Columbia University, USA; Arctic and North Pacific Oceans - hosted at Stockholm University, Sweden and the Center for Coastal and Ocean Mapping at the University of New Hampshire, USA.