 Catalogue PIGMA
Catalogue PIGMA
Benthic habitats
Type of resources
Available actions
Topics
Keywords
Contact for the resource
Provided by
Years
Formats
Representation types
status
Resolution
-
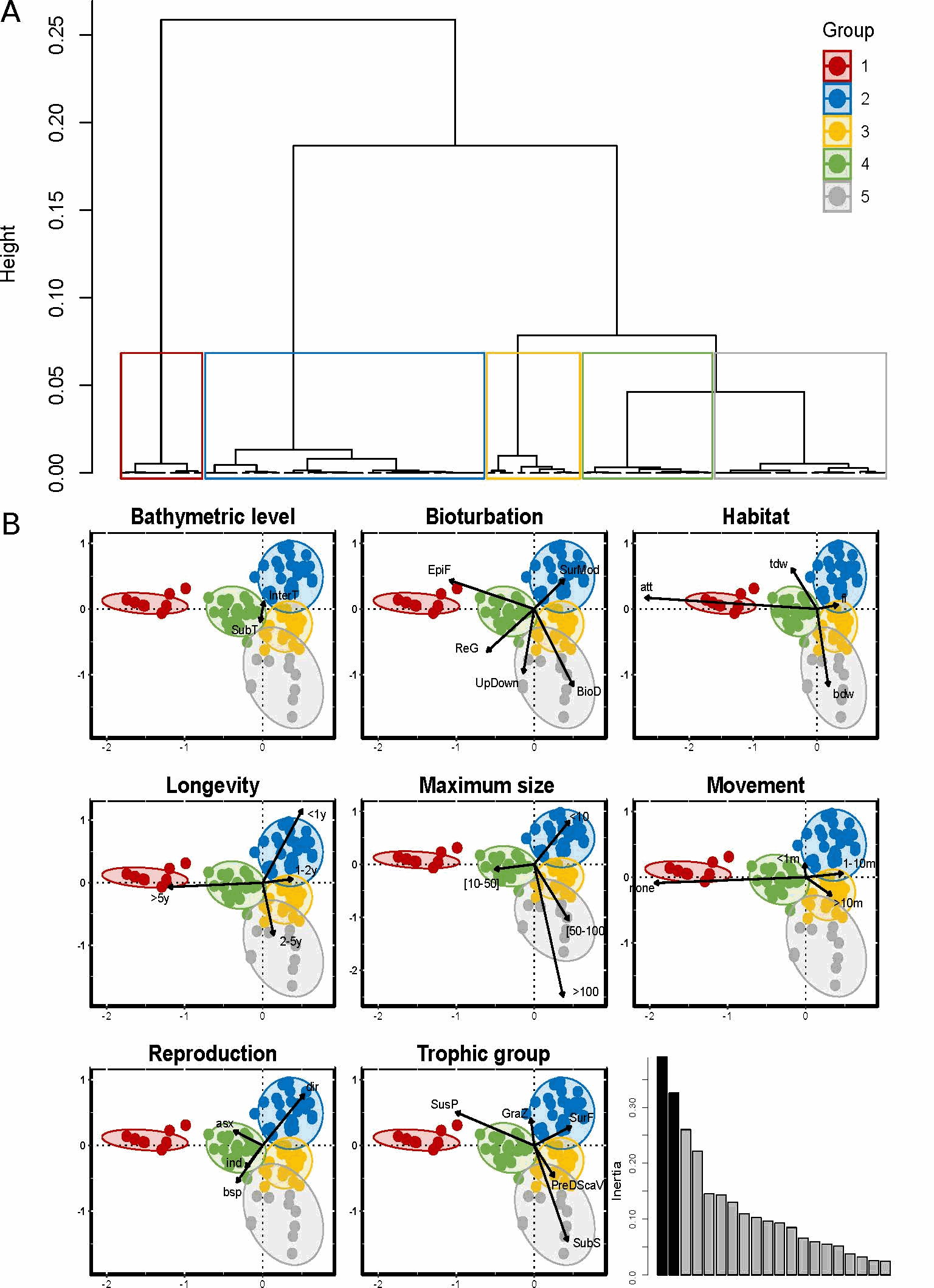
Reef-building species are recognized as having an important ecological role and as generally enhancing the diversity of benthic organisms in marine habitats. However, although these ecosystem engineers have a facilitating role for some species, they may exclude or compete with others. The honeycomb worm Sabellaria alveolata (Linnaeus, 1767) is an important foundation species, commonly found from northwest Ireland to northern Mauritania (Curd et al., 2020), whose reef structures increase the physical complexity of the marine benthos, supporting high levels of biodiversity. Local patterns and regional differences in taxonomic and functional diversity were examined in honeycomb worm reefs from ten sites along the northeastern Atlantic to explore variation in diversity across biogeographic regions and the potential effects of environmental drivers. To characterize the functional diversity at each site, a biological trait analysis (BTA) was conducted (Statzner et al., 1994). Here we present the functional trait database used for the benthic macrofauna found to live in association with honeycomb worm reefs. Eight biological traits (divided into 32 modalities) were selected (Table 1), providing information linked to the ecological functions performed by the associated macrofauna. The selected traits provide information on: (i) resource use and availability (by the trophic group of species, e.g. Thrush et al. 2006); (ii) secondary production and the amount of energy and organic matter (OM) produced based on the life cycle of the organisms (including longevity, maximum size and mode of reproduction, e.g. (Cusson and Bourget, 2005; Thrush et al., 2006) and; (iii) the behavior of the species in general [i.e. how these species occupy the environment and contribute to biogeochemical fluxes through habitat, movement, and bioturbation activity at different bathymetric levels, e.g. (Solan et al., 2004; Thrush et al., 2006; Queirós et al., 2013). Species were scored for each trait modality based on their affinity using a fuzzy coding approach (Chevenet et al., 1994), where multiple modalities can be attributed to a species if appropriate, and allowed for the incorporation of intraspecific variability in trait expression. The information concerning polychaetes was derived primarily from Fauchald et al (1979) and Jumars et al (2015). Information on other taxonomic groups was obtained either from databases of biological traits (www.marlin.ac.uk/biotic) or publications (Naylor, 1972; King, 1974; Caine, 1977; Lincoln, 1979; Holdich and Jones, 1983; Smaldon et al., 1993; Ingle, 1996; San Martín, 2003; Southward, 2008; Gil, 2011; Leblanc et al., 2011; Rumbold et al., 2012; San Martín and Worsfold, 2015; Jones et al., 2018). Map indicating the locations of the 10 study sites in the UK, France and Portugal within the four biogeographic provinces defined by Dinter (2001). (All sites were sampled in 8 different stations, except for UK4 where 5 stations were sampled).
-
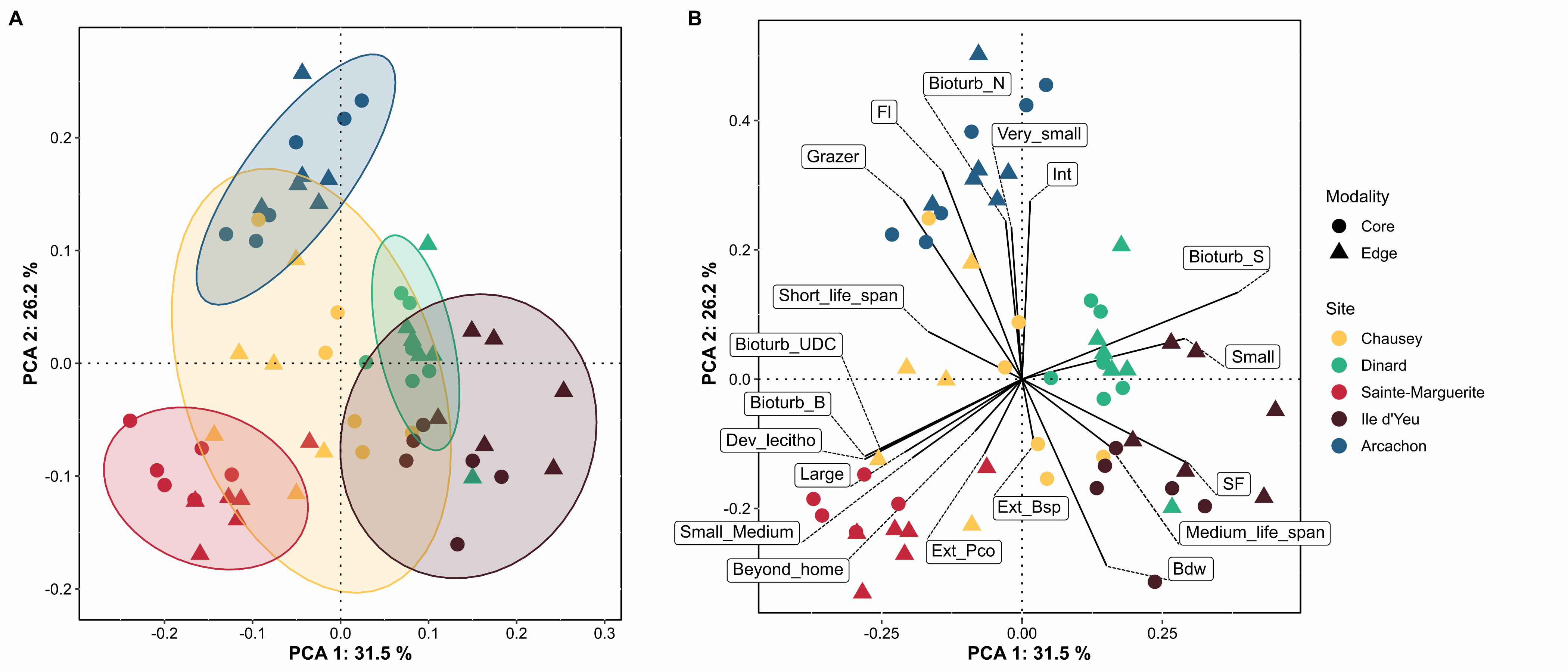
Zostera marina (Linnaeus, 1753) is a flowering marine plant that occurs from temperate to subantarctic regions (Green and Short, 2003), forming meadows that are recognized as being among the most important ecosystems on the planet (Costanza et al., 1997; Duffy, 2006; Duarte et al., 2008; Dewsbury et al., 2016). Eelgrass is a foundation species, providing essential functions and services including coastal protection, erosion control, nutrient cycling, water purification, carbon sequestration, as well as food and habitat for a variety of species (Duarte 2002; Heck et al. 2003; Healey & Hovel 2004, Orth et al. 2006; Barbier et al., 2011; Fourqurean et al. 2012; Cullen-Unsworth & Unsworth 2013; Schmidt et al. 2011, 2016). Eelgrass can have a strong influence on the spatial distribution of associated fauna by altering the hydrodynamics of the marine environment (Fonseca and Fisher 1986), stabilizing sediments (Orth et al. 2006), providing abundant resources, available surface area, and increased ecological niches. Meadows also provide protection from predation by providing greater habitat complexity both above and below ground (Heck and Wetstone 1977; Orth et al. 1984; Gartner et al. 2013, Reynolds et al., 2018). Local patterns and regional differences in the taxonomic and functional diversity of assemblages associated with five Zostera marina meadows occurring over a distance of 800 km along the coast of France were investigated with the objective of determining which factors control community composition within this habitat. To this end, we examined - and -diversity of species- and trait-based descriptors, focused on polychaetes; bivalves and gastropods, three diverse groups exhibiting a wide range of ecological strategies (Jumars, Dorgan, & Lindsay, 2015) and having central roles in ecosystem functioning through activities such as bioturbation or trophic regime (Queirós et al., 2013, Duffy et al., 2015). Here we present the abundance (Table 1) and the functional trait database (Table 2) used for the benthic macrofauna found to live in association with eelgrass meadows in Chausey, Dinard, Sainte-Marguerite, Ile d’Yeu and Arcachon, sampled in the fall of 2019. Eight biological traits (divided into 32 modalities, Table S1) were selected, providing information linked to the ecological functions performed by the associated macrofauna. The selected traits provide information on: (i) resource use and availability (by the trophic group of species, e.g. Thrush et al. 2006); (ii) secondary production and the amount of energy and organic matter (OM) produced based on the life cycle of the organisms (including longevity, maximum size and mode of reproduction, e.g. (Cusson and Bourget, 2005; Thrush et al., 2006) and; (iii) the behavior of the species in general [i.e. how these species occupy the environment and contribute to biogeochemical fluxes through habitat, movement, and bioturbation activity, e.g. (Solan et al., 2004; Thrush et al., 2006; Queirós et al., 2013). Species were scored for each trait modality based on their affinity using a fuzzy coding approach (Chevenet et al., 1994), where multiple modalities can be attributed to a species if appropriate, and allowed for the incorporation of intraspecific variability in trait expression. Information for polychaetes was primarily extracted from Fauchald et al (1979), Jumars et al (2015), and Boyé et al (2019). Information for mollusks was obtained either from biological trait databases (www.marlin.ac.uk/biotic, www.univie.ac.at/arctictraits, Bacouillard et al 2020) or from publications (e.g. Queiros et al. 2013; Thrush et al, 2006; Caine, 1977). Information was collected at the lowest possible taxonomic level and when missing was based on data available in other species of the genus, or in some cases, in the same family (only for traits with low variability for these families). Figure 1. Map indicating the locations of the 5 study sites of Zostera marina meadows in France: three in the the English Channel, and two in the Bay of Biscay (all sites were sampled in 6 different stations).
-

Numerous reef-forming species have declined dramatically in the last century, many of which have been insufficiently documented due to anecdotal or hard-to-access information. One of them, the honeycomb worm Sabellaria alveolata (L.) is a tube-building polychaete that can form large reefs, providing important ecosystem services such as coastal protection and habitat provision. It ranges from Scotland to Morocco, yet little is known about its distribution outside of the United Kingdom, where it is protected and where there is a strong heritage of natural history and sustained observations. As a result, online marine biodiversity information systems currently contain haphazardly distributed records of S. alveolata. One of the objectives of the REEHAB project (http://www.honeycombworms.org) was to combine historical records with contemporary data to document changes in the distribution and abundance of S. alveolata. Here we publish the result of the curation of 446 sources, gathered from literature, targeted surveys, local conservation reports, museum specimens, personal communications by authors and by their research teams, national biodiversity information systems (i.e. the UK National Biodiversity Network (NBN), https://nbn.org.uk/) and validated citizen science observations (i.e. https://www.inaturalist.org/). 80%[ar1] of these records were not previously referenced in any online information system. Additionally, historic field notebooks from Edouard Fischer-Piette and Gustave Gilson were scanned for S. alveolata information and manually entered. The original taxonomic identification of the 23296 S. alveolata records has been kept. Some identification errors may however be present, particularly in the English Channel and the North Sea where incorrectly identified observations of intertidal Sabellaria spinulosa were recorded. A further 229 observations are recorded as ‘Sabellaria spp.’ as the available information does not allow a species-level identification. Many sources reported abundances based on the semi-quantitative SACFOR scale while others simply noted its presence, and others still verified both its absence and presence. The result is a curated and comprehensive dataset spanning over two centuries on the past and present global distribution and abundance of S. alveolata. Sabellaria alveolata records projected onto a 50km grid. When SACFOR scale abundance scores were given to occurrence records, the highest abundance value per grid cell was retained.
-
French benthic invertebrates composition and abundance taxa data are collected during monitoring surveys on the English Channel / Bay of Biscay coasts and Mediterranean coast (Quadrige program code : REBENT_FAU, RSL_FAU). Protocols are implemented in the Water Framework Directive. Data are transmitted in a Seadatanet format (CDI + ODV) to EMODnet Biology european database. 498 ODV files have been generated from period 01/01/2003 to 31/12/2021.
-

The BenthOBS dataset includes long-term time series on marine benthic macrofauna, since 1967, along the whole French metropolitan coast. It includes 20 sampling location. BenthOBS aims to establish a national network for the observation of macrozoobenthos. In a context of global change, It is essential to have time series capable of highlighting and understanding ongoing changes in the specific diversity within communities and their consequences on the functioning of marine ecosystems. The BenthOBS network provides the scientific community and stackers with validated data on the following parameters: specific abundance, sediment size composition, sediment organic matter, sediment C content, sediment N content.
-

Particularly suited to the purpose of measuring the sensitivity of benthic communities to trawling, a trawl disturbance indicator (de Juan and Demestre, 2012, de Juan et al. 2009) was proposed based on benthic species life history traits to evaluate the sensibility of mega- and epifaunal community to fishing pressure known to have a physical impact on the seafloor (such as dredging and bottom trawling). The selected biological traits were chosen as they determine vulnerability to trawling: mobility, fragility, position on substrata, average size and feeding mode that can easily be related to the fragility, recoverability and vulnerability ecological concepts. Life history traits of species have been defined from the BIOTIC database (MARLIN, 2014) and from information given by Le Pape et al. (2007), Brindamour et al. (2009) and Garcia (2010). For missing life history traits, additional information from literature has been considered. The five categories retained are life history functional traits that were selected based on the knowledge of the response of benthic taxa to trawling disturbance (de Juan and Demestre, 2012). They reflect respectively the possibility to avoid direct gear impact, to benefit from trawling for feeding, to escape gear, to get caught by the net and to resist trawling/dredging action, each of these characteristics being either advantageous or sensitive to trawling. Then, to allow quantitative analysis, a score was assigned to each category: from low vulnerability (0) to high vulnerability (3). The five categories scores were then summed for each taxon (the highly vulnerable taxon could reach the maximum score is 15) and this value may be considered as a species index of sensitivity to trawling disturbance. The scores of 773 taxa commonly found in bottom trawl by-catch in the southern North Sea, English Channel and north-western Mediterranean were described.
-

Particularly suited to the purpose of measuring the sensitivity of benthic communities to trawling, a trawl disturbance indicator (de Juan and Demestre, 2012, de Juan et al. 2009) was proposed based on benthic species biological traits to evaluate the sensibility of mega- and epifaunal community to fishing pressure known to have a physical impact on the seafloor (such as dredging and bottom trawling). The selected biological traits were chosen as they determine vulnerability to trawling: mobility, fragility, position on substrata, average size and feeding mode that can easily be related to the fragility, recoverability and vulnerability ecological concepts. The five categories retained are functional traits that were selected based on the knowledge of the response of benthic taxa to trawling disturbance (de Juan et al., 2009). They reflect respectively the possibility to avoid direct gear impact, to benefit from trawling for feeding, to escape gear, to get caught by the net and to resist trawling/dredging action, each of these characteristics being either advantageous or sensitive to trawling. To expand this approach to that proposed by Certain et al. (2015), the protection status of certain species was also indicated. To enable quantitative analysis, a score was assigned to each category: from low sensitivity (0) to high sensitivity (3). Biological traits of species have been defined, from the BIOTIC database (MARLIN, 2014) and from information given by Garcia (2010), Le Pape et al. (2007) and Brind’Amour et al. (2009). For missing traits, additional information from literature has been considered. The protection status of each taxa was also scored: Atlantic species listed in OSPAR List of Threatened and/or Declining Species and Habitats (https://www.ospar.org/work-areas/bdc/species-habitats/list-of-threatened-declining-species-habitats) and Mediterranean species listed in Vulnerable Marine Ecosystems (FAO, 2018 and Oceana, 2017) were scored 3 and other species were scored 1. The scores of 1085 taxa commonly found in bottom trawl by-catch in the southern North Sea, English Channel and north-western Mediterranean were described.
-

Phenotypic plasticity, the ability of a single genotype to produce multiple phenotypes, is important for survival when species are faced with novel conditions. Theory predicts that range-edge populations will have greater phenotypic plasticity than core populations, but empirical examples from the wild are rare. The honeycomb worm, Sabellaria alveolata (L.), constructs the largest biogenic reefs in Europe, which support high biodiversity and numerous ecological functions. In order to assess the presence, causes and consequences of intraspecific variation in developmental plasticity and thermal adaptation in the honeycomb worm, we carried out common-garden experiments using the larvae of individuals sampled from along a latitudinal gradient covering the entire range of the species. We exposed larvae to three temperature treatments and measured phenotypic traits throughout development. We found phenotypic plasticity in larval growth rate but local adaptation in terms of larval period. The northern and southern range-edge populations of S. alveolata showed phenotypic plasticity for growth rate: growth rate increased as temperature treatment increased. In contrast, the core range populations showed no evidence of phenotypic plasticity. We present a rare case of range-edge plasticity at both the northern and southern range limit of species, likely caused by evolution of phenotypic plasticity during range expansion and its maintenance in highly heterogeneous environments. This dataset presents the raw image data collected for larval stages of Sabellaria alveolata from 5 populations across Europe and Northern Africa, exposed to 15, 20 and 25 C. Included are also opercular crown measurements used to estimate de size classes of individuals present in each population. All measurements made with the images collected are presented in an Excel spreadsheet, also available here.
-

The flat oyster Ostrea edulis is a European native species that once covered vast areas in the North Sea, on the Atlantic coast and in other European coastal waters including the Mediterranean region. All these populations have been heavily fished by dredging over the last three centuries. More recently, the emergence of parasites combined with the proliferation of various predators and many human-induced additional stressors have caused a dramatic decrease in the last remaining flat oyster populations. Today, this species has disappeared from many locations in Europe and is registered on the OSPAR (Oslo-Paris Convention for the Protection of the Marine environment of the North-East Atlantic) list of threatened and/or declining species (see https://www.ospar.org/work-areas/bdc/species-habitats/list-of-threatened-declining-species-habitats). In that context, since 2018, the Flat Oyster REcoVERy project (FOREVER) has been promoting the reestablishment of native oysters in Brittany (France). This multi-partner project, involving the CRC (Comité Régional de la Conchyliculture), IFREMER (Institut Français de Recherche pour l’Exploitation de la Mer), ESITC (École Supérieure d’Ingénieurs des Travaux de la Construction) Caen and Cochet Environnement, has consisted of (1) inventorying and evaluating the status of the main wild flat oyster populations across Brittany, (2) making detailed analysis of the two largest oyster beds in the bays of Brest and Quiberon to improve understanding of flat oyster ecology and recruitment variability and to suggest possible ways of improving recruitment, and (3) proposing practical measures for the management of wild beds in partnership with members of the shellfish industry and marine managers. the final report of this project is available on Archimer : https://doi.org/10.13155/79506. This survey is part of the task 1 of the FOREVER, which took place between 2017-2021. Some previous data, acquired with the same methodology and within the same geographic area have been also added to this dataset. These data were collected during 30 intertidal and diving surveys in various bays and inlets of the coast of Bretagne. The localization of these surveys has been guided by the help of historical maps. In the field, the methodology was simple enough to be easily implemented regardless of the configuration of the sampled site. The intertidal survey was conducted at very low tide (tidal range > 100) to sample the 0-1m level. Sampling was carried out randomly or systematically following the low water line. Where possible (in terms of visibility and accessibility), dive surveys were also carried out (0-10m depth), along 100m transects, using the same methodology of counting in a 1m2 quadrat. As often as possible, geo-referenced photographs were taken to show the appearance, density and habitat where Ostrea edulis was present. All these pictures are available in the image bank file. Overall, this dataset contains a total of 300 georeferenced records, where flat oysters have been observed. The dataset file contains also information concerning the surrounding habitat description and is organized according the OSPAR recommendations. This publication gives also a map, under a kml format showing each occurrence and its characteristics. This work was done in the framework of the following research project: " Inventaire, diagnostic écologique et restauration des principaux bancs d’huitres plates en Bretagne : le projet FOREVER. Contrat FEAMP 17/2215675".
-

Numerous reef-forming species have declined dramatically over the last century. Many of these declines have been insufficiently documented due to anecdotal or hard-to-access information. The Ross worm Sabellaria spinulosa (L.) is a tube-building polychaete that can form large mostly subtidal reefs, providing important ecosystem services such as coastal protection and habitat provision. It ranges from Scotland to Morocco and into the Mediterranean as far as the Adriatic, yet little is known about its distribution outside of the North & Wadden Seas, where it is protected under the OSPAR & HELCOM regional sea conventions respectively. As a result, online marine biodiversity information systems currently contain haphazardly distributed records of S. spinulosa. One of the objectives of the REEHAB project (http://www.honeycombworms.org) was to combine historical records with contemporary data to document changes in the distribution and abundance of the two Sabellaria species found in Europe, S. alveolata and S. spinulosa. Here we publish the result of the curation of 555 S. spinulosa sources, gathered from literature, targeted surveys, local conservation reports, museum specimens, personal communications by authors their research teams, national biodiversity information systems (i.e. the UK National Biodiversity Network (NBN), www.nbn.org.uk) and validated citizen science observations (i.e. https://www.inaturalist.org). 56% of these records were not previously referenced in any online information system. Additionally, historic samples from Gustave Gilson were scanned for S. spinulosa information and manually entered. The original taxonomic identification of the 40,261 S. spinulosa records has been kept. Some identification errors may however be present, particularly in the English Channel and Mediterranean where intertidal and shallow subtidal records can be mistaken for Sabellaria alveolata. A further 229 observations (16 sources) are recorded as ‘Sabellaria spp.’ as the available information did not provide an identification down to species level. Many sources reported abundances based on the semi-quantitative SACFOR scale whilst others simply noted its presence, and others still verified both its absence and presence. The result is a curated and comprehensive dataset spanning over two centuries on the past and present global distribution and abundance of S. spinulosa. Sabellaria spinulosa records projected onto a 50km grid. When SACFOR scale abundance scores were given to occurrence records, the highest abundance value per grid cell was retained.