 Catalogue PIGMA
Catalogue PIGMA
2025
Type of resources
Available actions
Topics
Keywords
Contact for the resource
Provided by
Years
Formats
Representation types
Update frequencies
status
Scale
Resolution
-
Micro litter - Density per net normalized per m³ - Research and monitoring protocols 2016/2023 v2025

This visualization product displays the density of floating micro-litter per net normalized per m³ per year from research and monitoring protocols. EMODnet Chemistry included the collection of marine litter in its 3rd phase. Before 2021, there was no coordinated effort at the regional or European scale for micro-litter. Given this situation, EMODnet Chemistry proposed to adopt the data gathering and data management approach as generally applied for marine data, i.e., populating metadata and data in the CDI Data Discovery and Access service using dedicated SeaDataNet data transport formats. EMODnet Chemistry is currently the official EU collector of micro-litter data from Marine Strategy Framework Directive (MSFD) National Monitoring activities (descriptor 10). A series of specific standard vocabularies or standard terms related to micro-litter have been added to SeaDataNet NVS (NERC Vocabulary Server) Common Vocabularies to describe the micro-litter. European micro-litter data are collected by the National Oceanographic Data Centres (NODCs). Micro-litter map products are generated from NODCs data after a test of the aggregated collection including data and data format checks and data harmonization. A filter is applied to represent only micro-litter sampled according to research and monitoring protocols as MSFD monitoring. Densities were calculated for each net using the following calculation: Density (number of particles per m³) = Micro-litter count / Sampling effort (m³) When the number of micro-litters was not filled, it was not possible to calculate the density. Percentiles 50, 75, 95 & 99 have been calculated taking into account data for all years. Warning: the absence of data on the map does not necessarily mean that they do not exist, but that no information has been entered in the National Oceanographic Data Centre (NODC) for this area.
-

This visualization product displays the density of floating micro-litter per net normalized in grams per km² per year from research and monitoring protocols. EMODnet Chemistry included the collection of marine litter in its 3rd phase. Before 2021, there was no coordinated effort at the regional or European scale for micro-litter. Given this situation, EMODnet Chemistry proposed to adopt the data gathering and data management approach as generally applied for marine data, i.e., populating metadata and data in the CDI Data Discovery and Access service using dedicated SeaDataNet data transport formats. EMODnet Chemistry is currently the official EU collector of micro-litter data from Marine Strategy Framework Directive (MSFD) National Monitoring activities (descriptor 10). A series of specific standard vocabularies or standard terms related to micro-litter have been added to SeaDataNet NVS (NERC Vocabulary Server) Common Vocabularies to describe the micro-litter. European micro-litter data are collected by the National Oceanographic Data Centres (NODCs). Micro-litter map products are generated from NODCs data after a test of the aggregated collection including data and data format checks and data harmonization. A filter is applied to represent only micro-litter sampled according to research and monitoring protocols as MSFD monitoring. Densities were calculated for each net using the following calculation: Density (weight of particles per km²) = Micro-litter weight / (Sampling effort (km) * Net opening (cm) * 0.00001) When information about the sampling effort (km) was lacking and point coordinates were known (start and end of the sampling), the sampling effort was calculated using the PostGIS ST_DistanceSpheroid function with a WGS84 measurement spheroid. When the weight of microlitters or the net opening was not filled, it was not possible to calculate the density. Percentiles 50, 75, 95 & 99 have been calculated taking into account data for all years. Warning: the absence of data on the map does not necessarily mean that they do not exist, but that no information has been entered in the National Oceanographic Data Centre (NODC) for this area.
-
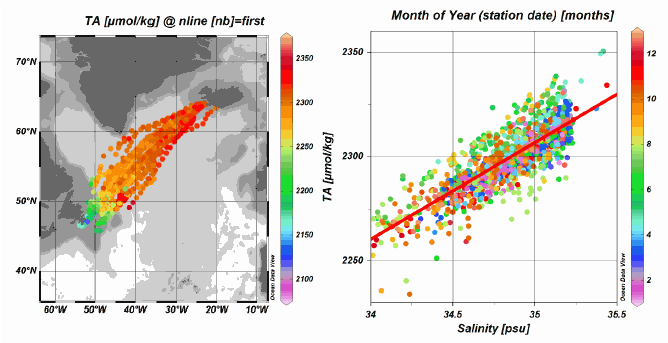
The SURATLANT dataset (SURveillance ATLANTique) consists of individual data of temperature, salinity, dissolved inorganic carbon (DIC) and its isotopic composition d13CDIC, total alkalinity (At), inorganic nutrients and water stable isotopes (δ18O and δD) collected mostly from ships of opportunity since 1993 along transects between Iceland and Newfoundland (shipping company EIMSKIP), as well as, since 2014, between west Greenland and Danemark (shipping company RAL). The data have been validated, qualified, and their accuracy and the overall characteristics of the data set are presented in a paper (Reverdin et al., 2018). The csv file provides a listing of the data with one line for each collection date. This includes collection date, position, temperature, salinity, and the measured, validated and in some cases adjusted variables, as well as a quality code following WOCE/GLODAP format. For water isotopes and isotopic composition of inroganic carbon, a code is also provided indicating the method of measurement used. An additional text-file provides a normalized average seasonal cycle of the 10 variables measured in 5 boxes between the vicinity of Newfoundland to the south-west of Iceland (corresponding to the figure 3 of the paper Reverdin et al in the References). Format and information is provided in the top 25 lines and the gridded seasonal cyle data start at line 26.
-

'''DEFINITION''' Important note to users: These data are not to be used for navigation. The data is 100 m resolution and as high quality as possible. It has been produced with state-of-the-art technology and validated to the best of the producer’s ability and where sufficient high-quality data were available. These data could be useful for planning and modelling purposes. The user should independently assess the adequacy of any material, data and/or information of the product before relying upon it. Neither Mercator Ocean International/Copernicus Marine Service nor the data originators are liable for any negative consequences following direct or indirect use of the product information, services, data products and/or data. Product overview: This is a satellite derived bathymetry product covering the global coastal area (where data retrieval is possible), with 100 m resolution, based on Sentinel-2. This global coastal product has been developed based on 3 methodologies: Intertidal Satellite-Derived Bathymetry; Physics-based optical Satellite-Derived Bathymetry from RTE inversion; and Wave Kinematics Satellite-Derived Bathymetry from wave dispersion. There is one dataset for each of the methods (including a quality index based on uncertainty) and an additional one where the three datasets were merged (also includes a quality index). Using their expertise and special techniques the consortium tried to achieve an optimal balance between coverage and data quality. '''DOI (product):''' https://doi.org/10.48670/mds-00364
-

EMODnet Chemistry aims to provide access to marine chemistry data sets and derived data products concerning eutrophication, ocean acidification, contaminants and litter. The chosen parameters are relevant for the Marine Strategy Framework Directive (MSFD), in particular for descriptors 5, 8, 9 and 10. The dataset contains standardized, harmonized and validated data collections from beach litter (monitoring and other sources). Datasets concerning beach and seafloor litter data are loaded in a central database after a semi-automated validation phase. Once loaded, a data assessment is performed in order to check data consistency and potential errors are corrected thanks to a feedback loop with data originators. For beach litter, the harmonized datasets contain all unrestricted EMODnet Chemistry data on beach litter, including monitoring data, data from cleaning surveys and data from research. A relevant part of the monitoring data has been considered for assessment purposes by the European institutions and therefore is tagged as MSFD_monitoring. EMODnet beach litter data and databases are hosted and maintained by 'Istituto Nazionale di Oceanografia e di Geofisica Sperimentale, Division of Oceanography (OGS/NODC)' from Italy. Data are formatted following Guidelines and forms for gathering marine litter data, which can be found at: https://doi.org/10.6092/15c0d34c-a01a-4091-91ac-7c4f561ab508 The updated vocabularies of admitted values are available at: https://nodc.ogs.it/marinelitter/vocab The harmonized datasets can be downloaded as EMODnet Beach litter data format Version 7.0, which is a spreadsheet file composed of 4 sheets: beach metadata, survey metadata, animals and litter. Local_CDI field in the survey metadata sheet allows to retrieve the original data.
-
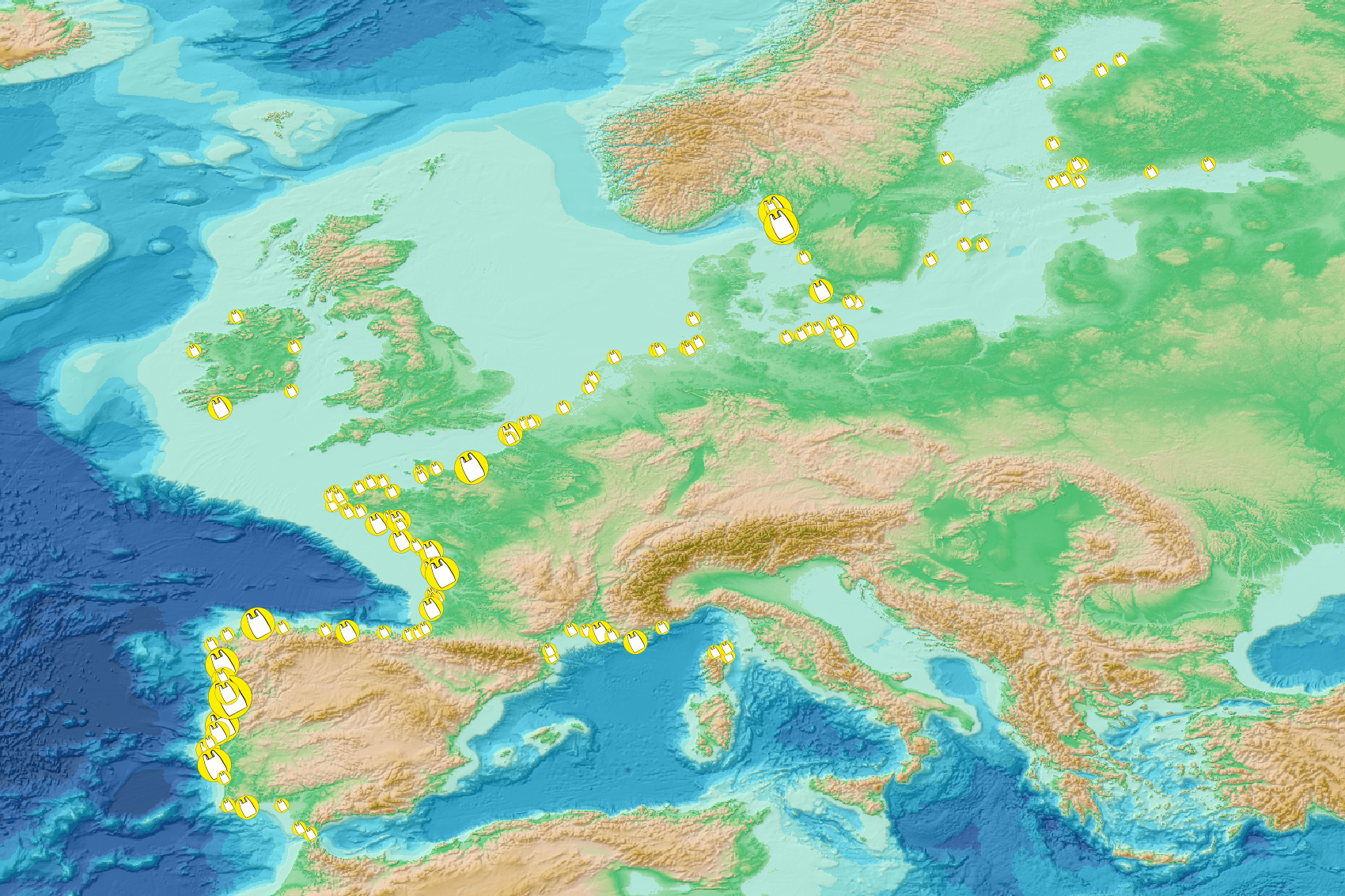
This visualization product displays the fishing & aquaculture related plastic items abundance of marine macro-litter (> 2.5cm) per beach per year from Marine Strategy Framework Directive (MSFD) monitoring surveys. EMODnet Chemistry included the collection of marine litter in its 3rd phase. Since the beginning of 2018, data of beach litter have been gathered and processed in the EMODnet Chemistry Marine Litter Database (MLDB). The harmonization of all the data has been the most challenging task considering the heterogeneity of the data sources, sampling protocols and reference lists used on a European scale. Preliminary processings were necessary to harmonize all the data: - Exclusion of OSPAR 1000 protocol: in order to follow the approach of OSPAR that it is not including these data anymore in the monitoring; - Selection of MSFD surveys only (exclusion of other monitoring, cleaning and research operations); - Exclusion of beaches without coordinates; - Selection of plastic bags related items only. The list of selected items is attached to this metadata. This list was created using EU Marine Beach Litter Baselines, the European Threshold Value for Macro Litter on Coastlines and the Joint list of litter categories for marine macro-litter monitoring from JRC (these three documents are attached to this metadata); - Normalization of survey lengths to 100m & 1 survey / year: in some case, the survey length was not exactly 100m, so in order to be able to compare the abundance of litter from different beaches a normalization is applied using this formula: Number of plastic bags related items of the survey (normalized by 100 m) = Number of plastic bags related items of the survey x (100 / survey length) Then, this normalized number of plastic bags related items is summed to obtain the total normalized number of plastic bags related items for each survey. Finally, the median abundance of plastic bags related items for each beach and year is calculated from these normalized abundances of plastic bags related items per survey. Sometimes the survey length was null or equal to 0. Assuming that the MSFD protocol has been applied, the length has been set at 100m in these cases. Percentiles 50, 75, 95 & 99 have been calculated taking into account plastic bags related items from MSFD data for all years. More information is available in the attached documents. Warning: the absence of data on the map does not necessarily mean that they do not exist, but that no information has been entered in the Marine Litter Database for this area.
-

The rasters correspond to the prediction uncertainties associted with the production of Mediterranean bioregions of megabenthic communities
-

This visualization product displays the cigarette related items abundance of marine macro-litter (> 2.5cm) per beach per year from Marine Strategy Framework Directive (MSFD) monitoring surveys without UNEP-MARLIN data. EMODnet Chemistry included the collection of marine litter in its 3rd phase. Since the beginning of 2018, data of beach litter have been gathered and processed in the EMODnet Chemistry Marine Litter Database (MLDB). The harmonization of all the data has been the most challenging task considering the heterogeneity of the data sources, sampling protocols and reference lists used on a European scale. Preliminary processings were necessary to harmonize all the data: - Exclusion of OSPAR 1000 protocol: in order to follow the approach of OSPAR that it is not including these data anymore in the monitoring; - Selection of MSFD surveys only (exclusion of other monitoring, cleaning and research operations); - Exclusion of beaches without coordinates; - Selection of cigarette related items only. The list of selected items is attached to this metadata. This list was created using EU Marine Beach Litter Baselines, the European Threshold Value for Macro Litter on Coastlines and the Joint list of litter categories for marine macro-litter monitoring from JRC (these three documents are attached to this metadata); - Selection of surveys referring to the UNEP-MARLIN list: the UNEP-MARLIN protocol differs from the other types of monitoring in that cigarette butts are surveyed in a 10m square. To avoid comparing abundances from very different protocols, the choice has been made to distinguish in two maps the cigarette related items results associated with the UNEP-MARLIN list from the others; - Normalization of survey lengths to 100m & 1 survey / year: in some case, the survey length was not exactly 100m, so in order to be able to compare the abundance of litter from different beaches a normalization is applied using this formula: Number of cigarette related items of the survey (normalized by 100 m) = Number of cigarette related items of the survey x (100 / survey length) Then, this normalized number of cigarette related items is summed to obtain the total normalized number of cigarette related items for each survey. Finally, the median abundance of cigarette related items for each beach and year is calculated from these normalized abundances of cigarette related items per survey. Sometimes the survey length was null or equal to 0. Assuming that the MSFD protocol has been applied, the length has been set at 100m in these cases. Percentiles 50, 75, 95 & 99 have been calculated taking into account cigarette related items from MSFD monitoring data (excluding UNEP-MARLIN protocol) for all years. More information is available in the attached documents. Warning: the absence of data on the map does not necessarily mean that they do not exist, but that no information has been entered in the Marine Litter Database for this area.
-
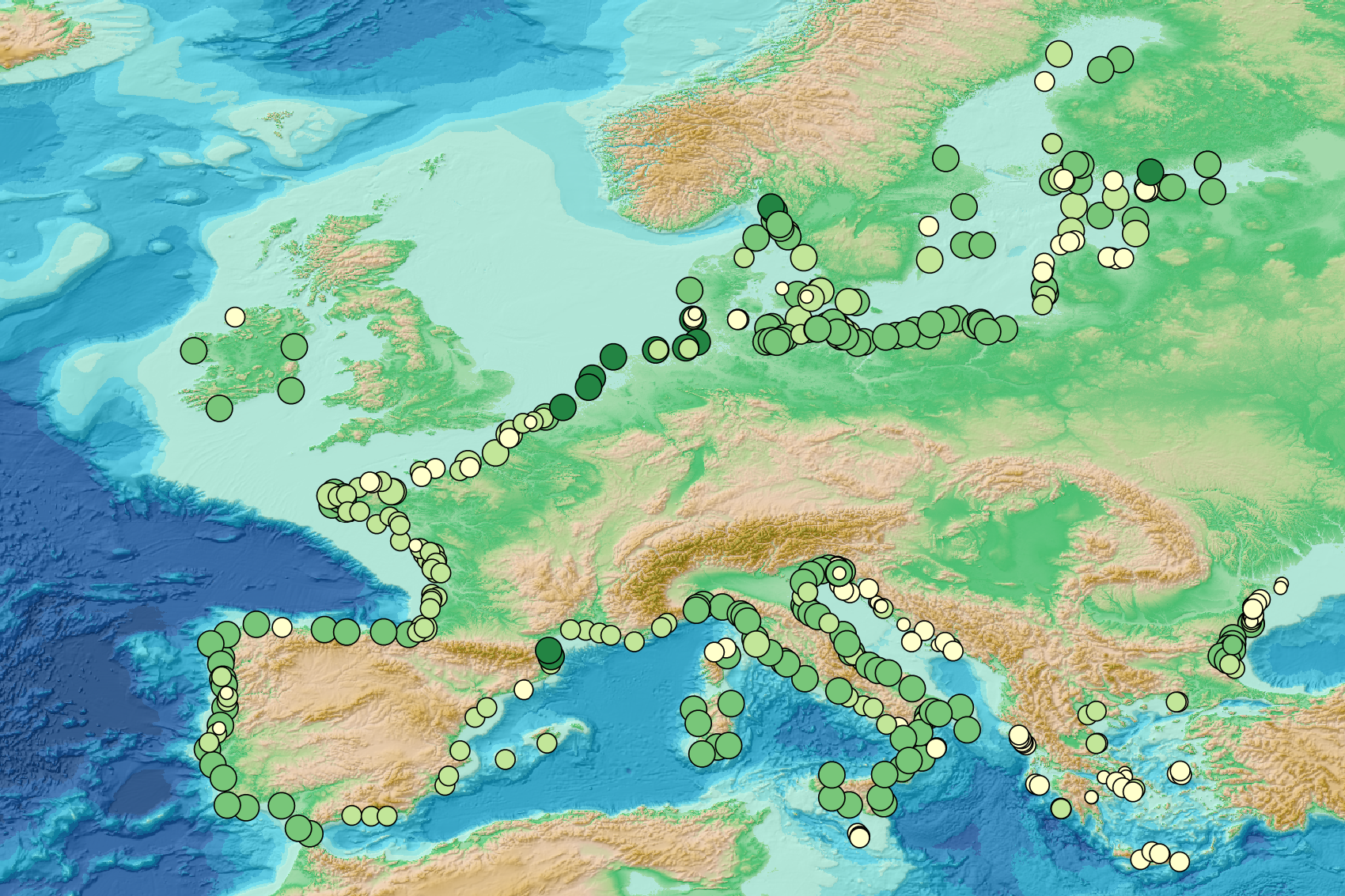
This visualization product displays the number of Marine Strategy Framework Directive (MSFD) monitoring surveys and the associated temporal coverage per beach. EMODnet Chemistry included the collection of marine litter in its 3rd phase. Since the beginning of 2018, data of beach litter have been gathered and processed in the EMODnet Chemistry Marine Litter Database (MLDB). The harmonization of all the data has been the most challenging task considering the heterogeneity of the data sources, sampling protocols and reference lists used on a European scale. Preliminary processings were necessary to harmonize all the data: - Exclusion of OSPAR 1000 protocol: in order to follow the approach of OSPAR that it is not including these data anymore in the monitoring; - Selection of MSFD surveys only (exclusion of other monitoring, cleaning and research operations); - Exclusion of beaches without coordinates. More information is available in the attached documents. Warning: the absence of data on the map does not necessarily mean that they do not exist, but that no information has been entered in the Marine Litter Database for this area.
-

Rapid changes in ocean circulation and climate have been observed in marine-sediment and ice cores over the last glacial period and deglaciation, highlighting the non-linear character of the climate system and underlining the possibility of rapid climate shifts in response to anthropogenic greenhouse gas forcing. To date, these rapid changes in climate and ocean circulation are still not fully explained. One obstacle hindering progress in our understanding of the interactions between past ocean circulation and climate changes is the difficulty of accurately dating marine cores. Here, we present a set of 92 marine sediment cores from the Atlantic Ocean for which we have established age-depth models that are consistent with the Greenland GICC05 ice core chronology, and computed the associated dating uncertainties, using a new deposition modeling technique. This is the first set of consistently dated marine sediment cores enabling paleoclimate scientists to evaluate leads/lags between circulation and climate changes over vast regions of the Atlantic Ocean. Moreover, this data set is of direct use in paleoclimate modeling studies.