 Catalogue PIGMA
Catalogue PIGMA
Analysis data
Type of resources
Available actions
Topics
Keywords
Contact for the resource
Provided by
Years
Formats
Representation types
-
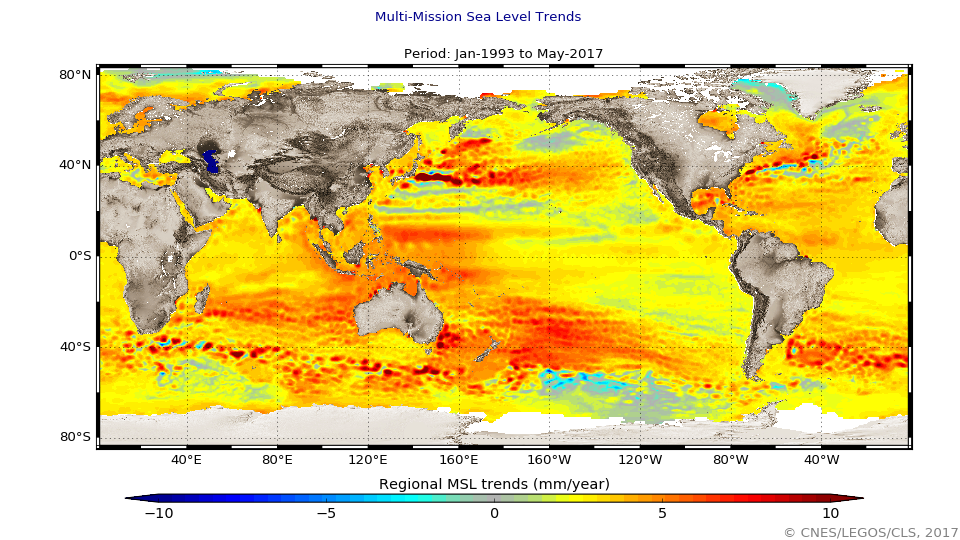
The global mean level of the oceans is one of the most important indicators of climate change. It incorporates the reactions from several different components of the climate system. Precise monitoring of changes in the mean level of the oceans, particularly through the use of altimetry satellites, is vitally important, for understanding not just the climate but also the socioeconomic consequences of any rise in sea level.
-

Numerous reef-forming species have declined dramatically in the last century, many of which have been insufficiently documented due to anecdotal or hard-to-access information. One of them, the honeycomb worm Sabellaria alveolata (L.) is a tube-building polychaete that can form large reefs, providing important ecosystem services such as coastal protection and habitat provision. It ranges from Scotland to Morocco, yet little is known about its distribution outside of the United Kingdom, where it is protected and where there is a strong heritage of natural history and sustained observations. As a result, online marine biodiversity information systems currently contain haphazardly distributed records of S. alveolata. One of the objectives of the REEHAB project (http://www.honeycombworms.org) was to combine historical records with contemporary data to document changes in the distribution and abundance of S. alveolata. Here we publish the result of the curation of 331 sources, gathered from literature, targeted surveys, local conservation reports, museum specimens, personal communications by authors and by their research teams, national biodiversity information systems (i.e. the UK National Biodiversity Network (NBN), https://nbn.org.uk/) and validated citizen science observations (i.e. https://www.inaturalist.org/). 80% of these records were not previously referenced in any online information system. Additionally, historic field notebooks from Edouard Fischer-Piette and Gustave Gilson were scanned for S. alveolata information and manually entered. Each of the 21512 S. alveolata records were checked for spatial and taxonomic accuracy, particularly in the English Channel and the North Sea where incorrectly identified observations of intertidal Sabellaria spinulosa were recorded. A further 54 observations are recorded as ‘Sabellaria spp.’ as the available information did not allow for an identification to species level. Many sources reported abundances based on the semi-quantitative SACFOR scale whilst others simply noted its presence, and others still verified both its absence and presence. The result is a curated and comprehensive dataset spanning over two centuries on the past and present global distribution and abundance of S. alveolata. Sabellaria alveolata records projected onto a 50km grid. When SACFOR scale abundance scores were given to occurrence records, the highest abundance value per grid cell was retained.
-
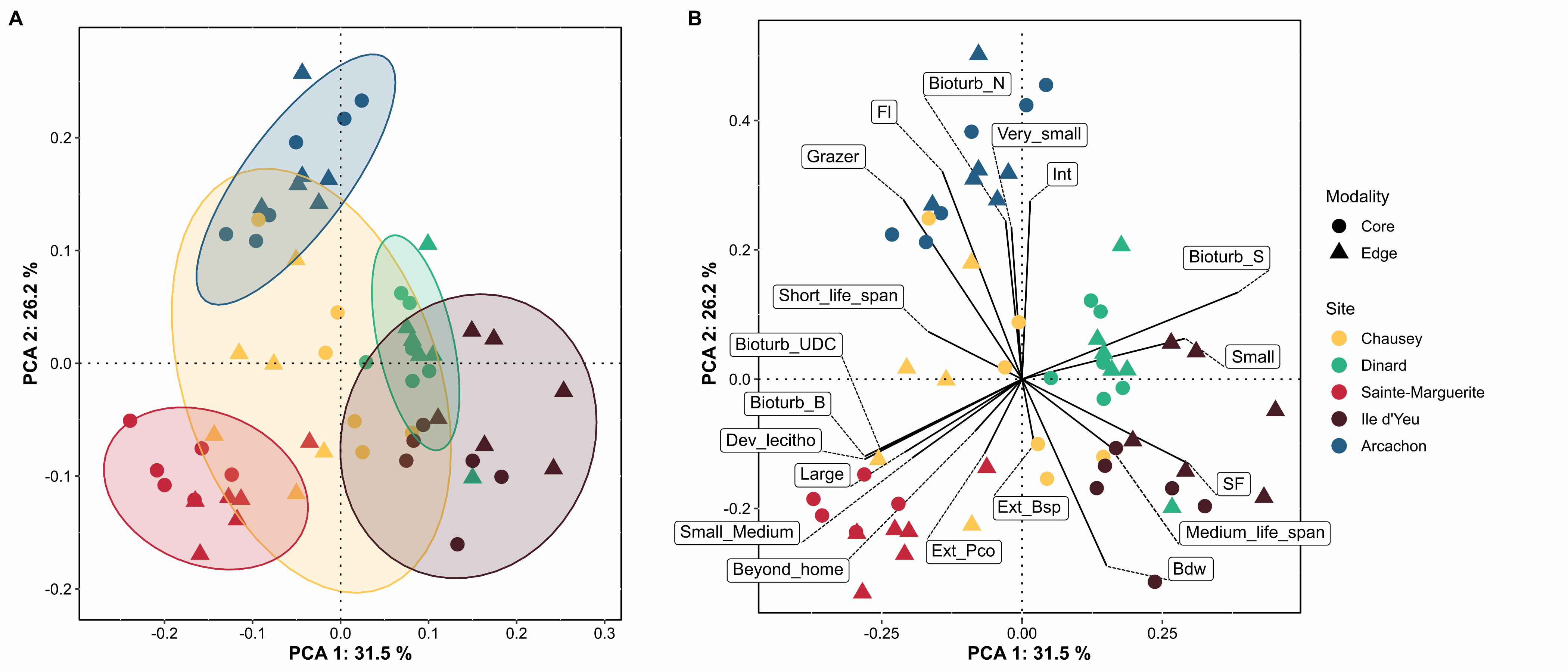
Zostera marina (Linnaeus, 1753) is a flowering marine plant that occurs from temperate to subantarctic regions (Green and Short, 2003), forming meadows that are recognized as being among the most important ecosystems on the planet (Costanza et al., 1997; Duffy, 2006; Duarte et al., 2008; Dewsbury et al., 2016). Eelgrass is a foundation species, providing essential functions and services including coastal protection, erosion control, nutrient cycling, water purification, carbon sequestration, as well as food and habitat for a variety of species (Duarte 2002; Heck et al. 2003; Healey & Hovel 2004, Orth et al. 2006; Barbier et al., 2011; Fourqurean et al. 2012; Cullen-Unsworth & Unsworth 2013; Schmidt et al. 2011, 2016). Eelgrass can have a strong influence on the spatial distribution of associated fauna by altering the hydrodynamics of the marine environment (Fonseca and Fisher 1986), stabilizing sediments (Orth et al. 2006), providing abundant resources, available surface area, and increased ecological niches. Meadows also provide protection from predation by providing greater habitat complexity both above and below ground (Heck and Wetstone 1977; Orth et al. 1984; Gartner et al. 2013, Reynolds et al., 2018). Local patterns and regional differences in the taxonomic and functional diversity of assemblages associated with five Zostera marina meadows occurring over a distance of 800 km along the coast of France were investigated with the objective of determining which factors control community composition within this habitat. To this end, we examined - and -diversity of species- and trait-based descriptors, focused on polychaetes; bivalves and gastropods, three diverse groups exhibiting a wide range of ecological strategies (Jumars, Dorgan, & Lindsay, 2015) and having central roles in ecosystem functioning through activities such as bioturbation or trophic regime (Queirós et al., 2013, Duffy et al., 2015). Here we present the abundance (Table 1) and the functional trait database (Table 2) used for the benthic macrofauna found to live in association with eelgrass meadows in Chausey, Dinard, Sainte-Marguerite, Ile d’Yeu and Arcachon, sampled in the fall of 2019. Eight biological traits (divided into 32 modalities, Table S1) were selected, providing information linked to the ecological functions performed by the associated macrofauna. The selected traits provide information on: (i) resource use and availability (by the trophic group of species, e.g. Thrush et al. 2006); (ii) secondary production and the amount of energy and organic matter (OM) produced based on the life cycle of the organisms (including longevity, maximum size and mode of reproduction, e.g. (Cusson and Bourget, 2005; Thrush et al., 2006) and; (iii) the behavior of the species in general [i.e. how these species occupy the environment and contribute to biogeochemical fluxes through habitat, movement, and bioturbation activity, e.g. (Solan et al., 2004; Thrush et al., 2006; Queirós et al., 2013). Species were scored for each trait modality based on their affinity using a fuzzy coding approach (Chevenet et al., 1994), where multiple modalities can be attributed to a species if appropriate, and allowed for the incorporation of intraspecific variability in trait expression. Information for polychaetes was primarily extracted from Fauchald et al (1979), Jumars et al (2015), and Boyé et al (2019). Information for mollusks was obtained either from biological trait databases (www.marlin.ac.uk/biotic, www.univie.ac.at/arctictraits, Bacouillard et al 2020) or from publications (e.g. Queiros et al. 2013; Thrush et al, 2006; Caine, 1977). Information was collected at the lowest possible taxonomic level and when missing was based on data available in other species of the genus, or in some cases, in the same family (only for traits with low variability for these families). Figure 1. Map indicating the locations of the 5 study sites of Zostera marina meadows in France: three in the the English Channel, and two in the Bay of Biscay (all sites were sampled in 6 different stations).
-
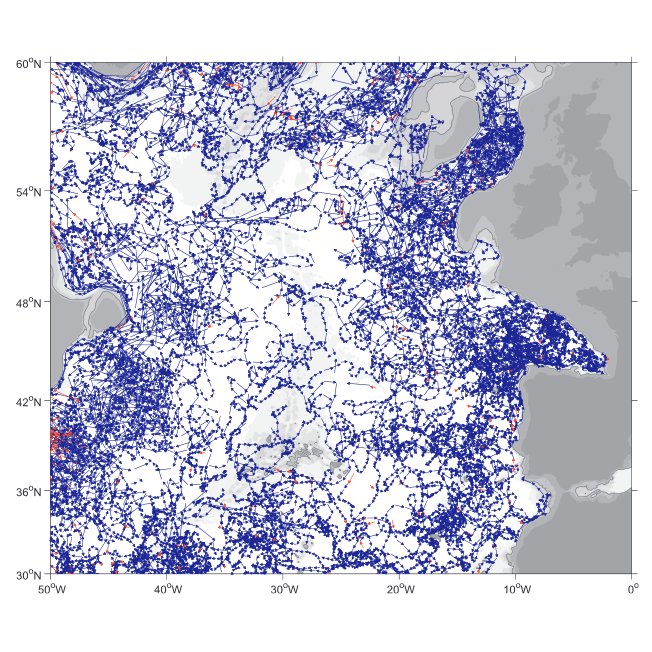
A world deep displacement dataset comprising more than 1600 000 Argo floats deep displacements, has been produced from the global Argo float database (GDAC). ANDRO dataset was completed over the period 2000-2009, then was partially but yearly updated since 2010. ANDRO actual contents and format is described in the user guide, which must be carefully read before using ANDRO (ANDRO format is also described in Ollitrault M. et al (2013)). One important feature of ANDRO is that the pressures measured during float drifts at depth, and suitably averaged are preserved in ANDRO (see Figure 2). To reach this goal, it was necessary to reprocess most of the Argo raw data, because of the many different decoding versions (roughly 100) not always applied by the DACs to the displacement data because they were mainly interested in the p,t,S profiles. The result of our work was the production of comprehensive files, named DEP (for déplacements in French), containing all the possibly retrievable float data. For detailed information and status of the last released ANDRO product, please visit the dedicated Argo France web page: https://www.umr-lops.fr/SNO-Argo/Products/ANDRO-Argo-floats-displacements-Atlas
-
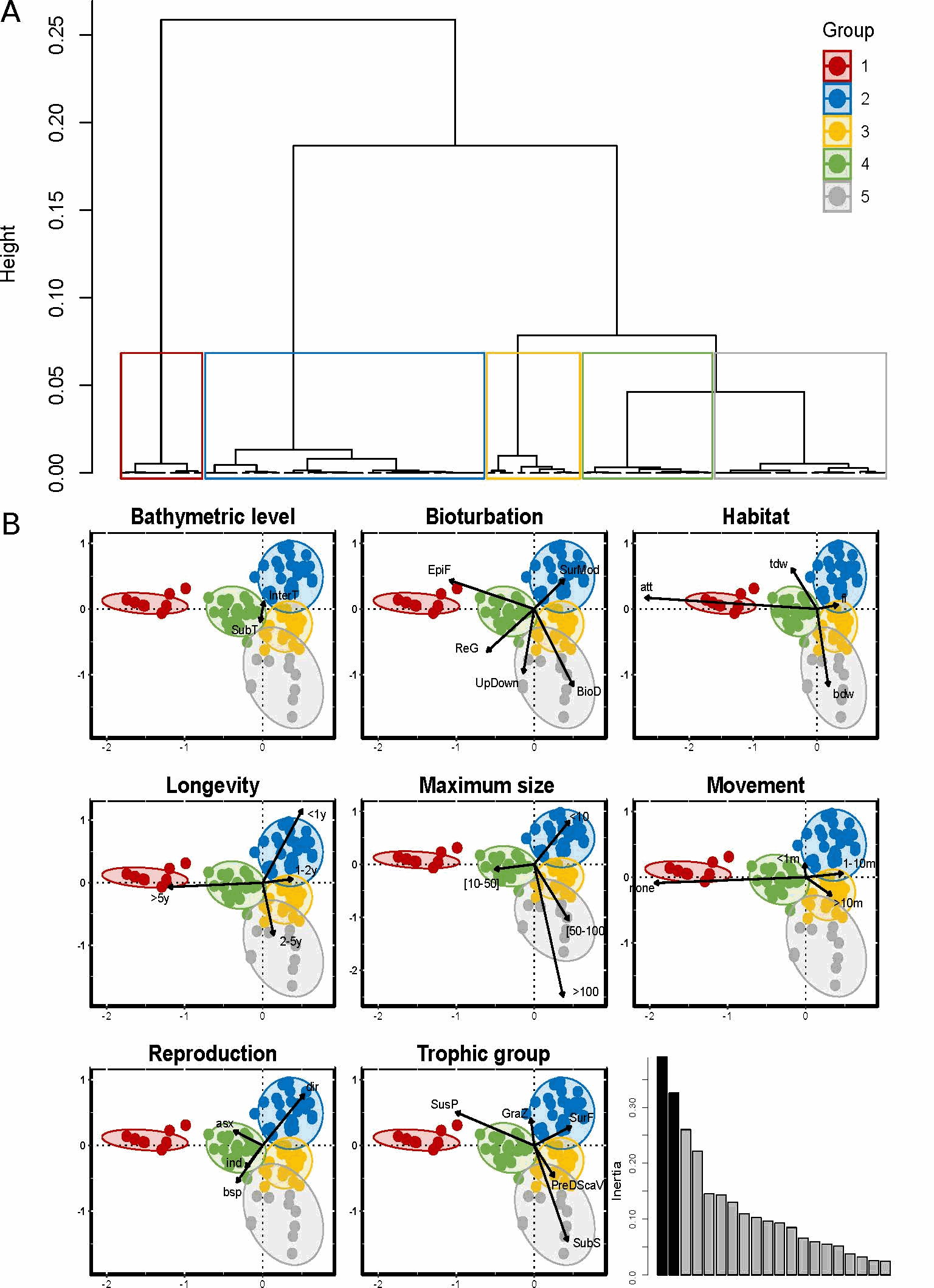
Reef-building species are recognized as having an important ecological role and as generally enhancing the diversity of benthic organisms in marine habitats. However, although these ecosystem engineers have a facilitating role for some species, they may exclude or compete with others. The honeycomb worm Sabellaria alveolata (Linnaeus, 1767) is an important foundation species, commonly found from northwest Ireland to northern Mauritania (Curd et al., 2020), whose reef structures increase the physical complexity of the marine benthos, supporting high levels of biodiversity. Local patterns and regional differences in taxonomic and functional diversity were examined in honeycomb worm reefs from ten sites along the northeastern Atlantic to explore variation in diversity across biogeographic regions and the potential effects of environmental drivers. To characterize the functional diversity at each site, a biological trait analysis (BTA) was conducted (Statzner et al., 1994). Here we present the functional trait database used for the benthic macrofauna found to live in association with honeycomb worm reefs. Eight biological traits (divided into 32 modalities) were selected (Table 1), providing information linked to the ecological functions performed by the associated macrofauna. The selected traits provide information on: (i) resource use and availability (by the trophic group of species, e.g. Thrush et al. 2006); (ii) secondary production and the amount of energy and organic matter (OM) produced based on the life cycle of the organisms (including longevity, maximum size and mode of reproduction, e.g. (Cusson and Bourget, 2005; Thrush et al., 2006) and; (iii) the behavior of the species in general [i.e. how these species occupy the environment and contribute to biogeochemical fluxes through habitat, movement, and bioturbation activity at different bathymetric levels, e.g. (Solan et al., 2004; Thrush et al., 2006; Queirós et al., 2013). Species were scored for each trait modality based on their affinity using a fuzzy coding approach (Chevenet et al., 1994), where multiple modalities can be attributed to a species if appropriate, and allowed for the incorporation of intraspecific variability in trait expression. The information concerning polychaetes was derived primarily from Fauchald et al (1979) and Jumars et al (2015). Information on other taxonomic groups was obtained either from databases of biological traits (www.marlin.ac.uk/biotic) or publications (Naylor, 1972; King, 1974; Caine, 1977; Lincoln, 1979; Holdich and Jones, 1983; Smaldon et al., 1993; Ingle, 1996; San Martín, 2003; Southward, 2008; Gil, 2011; Leblanc et al., 2011; Rumbold et al., 2012; San Martín and Worsfold, 2015; Jones et al., 2018). Map indicating the locations of the 10 study sites in the UK, France and Portugal within the four biogeographic provinces defined by Dinter (2001). (All sites were sampled in 8 different stations, except for UK4 where 5 stations were sampled).
-

Good Environmental Status assessment (GES) for descriptor 8 (contaminants, D8) of the Marine Strategy Framework Directive (MSFD) is reached when concentrations of contaminants are at levels not giving rise to pollution effects. It is described by 4 criteria among which the first one focus on the concentration of the contaminants in the environment (criteria 1 of the D8, D8C1). The environmental status for D8 in France includes assessment of contaminant concentrations in sediment, bivalves, fish, birds, mammals to cover the French marine area the continental shelf from the coast line). The 8 tables below present the assessment of the chemical contamination in sediment and bivalves on the coastal area of the 4 French marine subregions for D8 as part of the 2024 GES assessment. These tables report the status and temporal trends of each station x matrice x substance triplet in each of the 4 French marine subregions. Explanation on how to read the cells is given in the “read file”. The environmental assessment for D8 in France can be found in Mauffret al., 2023 (DOI:10.13155/97214). It includes 17 national indicator assessments, 4 OSPAR indicators and integrated assessment in selected assessment units at the level of the criteria 1 and 2.
-
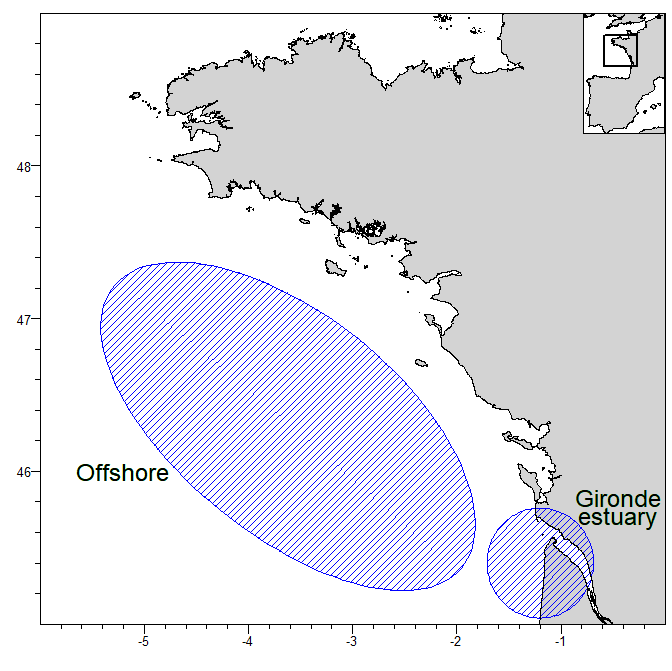
Input data, TMB (C++) and R code for close-kin mark recapture (CKMR) abundance estimation for two subpopulations of thornback ray (Raja clavata) in the central Bay of Biscay (offshore and Gironde estuary, see figure). Samples were taken between 2015 to 2020. Parent-offspring pairs were identified using SNP genotype information. Birth years of individuals were estimated using length information and growth curves by sex.
-
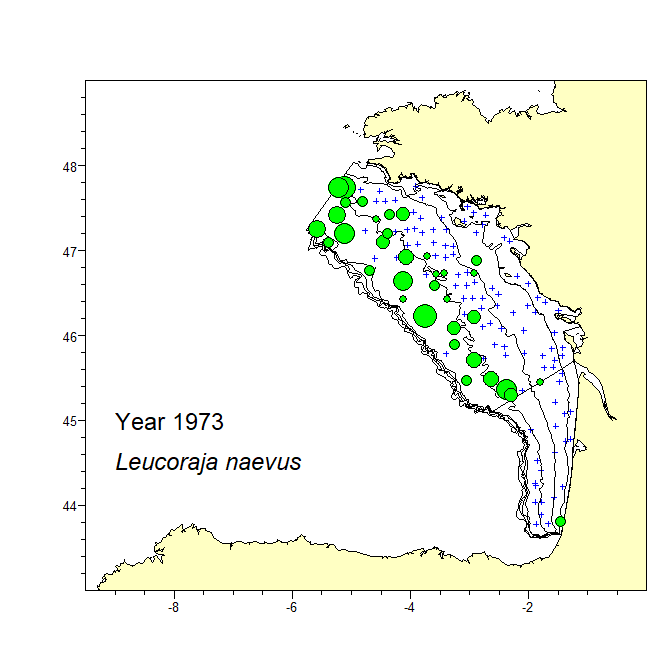
A consistent dataset of bottom trawl survey data spanning 47 years in the Bay of Biscay was assembled. The dataset includes data from the current EVHOE survey from 1987 to 2019 and two previous surveys carried out in 1973 and 1976. The recent EVHOE time-series from 1997 is also available from DATRAS (https://www.ices.dk/data/data-portals/Pages/DATRAS.aspx). The catch in numbers and weight (kg) per haul of all Rajiformes species caught in these surveys is provided. Haul information is provided for all hauls, including those with no catch of Rajiformes. Areas of the sampling strata of the survey and spatial polygones of these strata are provided in separate files.
-
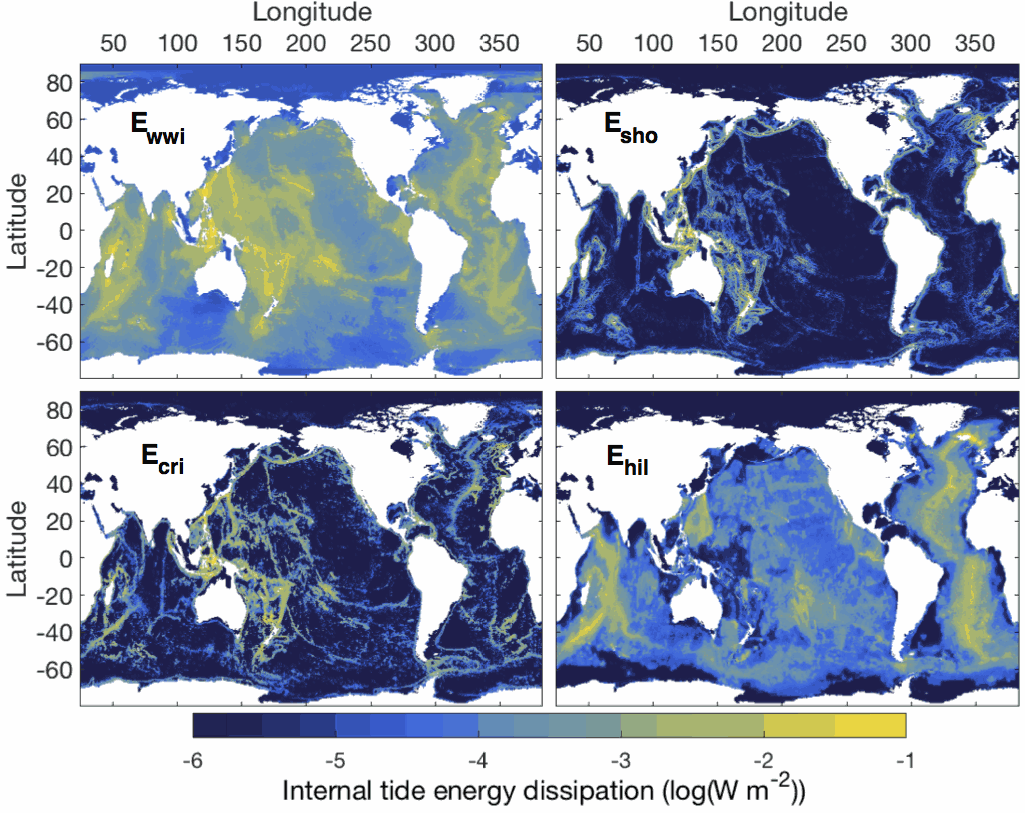
This dataset comprises two netcdf files. The first file contains the six global two-dimensional maps necessary to implement the tidal mixing parameterization presented in de Lavergne et al. (2020). Four power fields (E_wwi, E_sho, E_cri and E_hil) represent depth-integrated internal tide energy dissipation, with units of Watts per square meter. Each power field corresponds to a specific dissipative process and associated vertical structure of turbulence production. The two remaining fields, H_cri and H_bot, are decay heights (with units of meters) that enter the vertical structures of the E_cri and E_hil components, respectively. The second file contains three-dimensional fields of turbulence production (with units of Watts per kilogram) obtained by application of the parameterization to the WOCE global hydrographic climatology. The file includes the total turbulence production (epsilon_tid), its four components (epsilon_wwi, epsilon_sho, epsilon_cri, epsilon_hil), and the underlying hydrographic fields, as a function of longitude, latitude and depth. All maps have a horizontal resolution of 0.5º. Detailed documentation of the parameterization can be found in the following publication: de Lavergne, C., Vic, C., Madec, G., Roquet, F., Waterhouse, A.F., Whalen, C.B., Cuypers, Y., Bouruet-Aubertot, P., Ferron, B., Hibiya, T. A parameterization of local and remote tidal mixing. Journal of Advances in Modeling Earth Systems, 12, e2020MS002065 (2020). https://doi.org/10.1029/2020MS002065
-
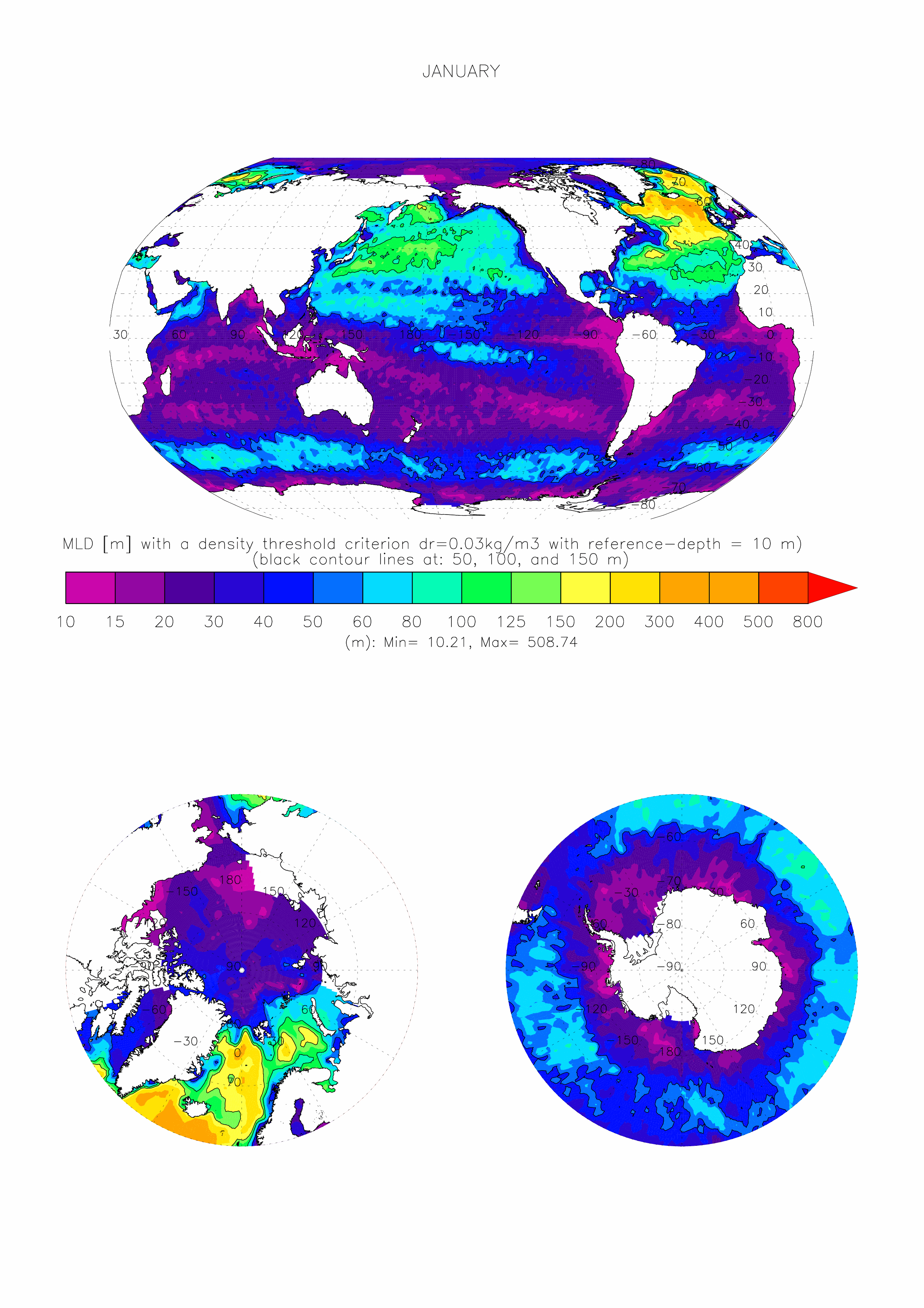
The dataset made available here is the monthly climatology (i.e. 12 months) of ocean surface Mixed Layer Depth (MLD) over the global ocean, at 1 degree x 1 degree spatial resolution. The climatology is based on about 7.3 million casts/profiles of temperature and salinity measurements made at sea between January 1970 and December 2021. Those profiles data come from the ARGO program and from the NCEI-NOAA World Ocean Database (WOD, Boyer et al. 2018). The MLD is computed on each individual cast/profile using a threshold criterion. The depth of the mixed layer is defined as the shallowest depth where the surface potential density of the profile is superior to a reference value taken close to the surface added with the chosen threshold. Here we take a threshold value for the density of 0.03kg/m3, and a surface reference depth fixed at 10m (de Boyer Montégut et al., 2004). This mixed layer is by definition homogeneous in density (up to 0.03 kg/m3 variations) and can also be called an isopycnal layer. It is especially intended for validation of MLD fields of the Ocean General Circulation Models of the ocean sciences community (e.g. Tréguier et al., 2023, Iovino et al. 2023, using v2022 of this dataset). More information and some other related datasets can be found at : https://cerweb.ifremer.fr/mld (or https://www.umr-lops.fr/en/Data/MLD redirecting to previous page).